2019-09-10 基礎生物学研究所,名古屋大学,高知大学
季節ごとに見られる色とりどりの花の形成は、私たちが身近に感じる自然現象の一つです。植物では良く知られている花芽の形成ですが、生物の進化過程においてどのように獲得されたのかはわかっていませんでした。基礎生物学研究所 環境光生物学研究部門の 得津 隆太郎 助教、皆川 純 教授らは、名古屋大学遺伝子実験施設の 松尾 拓哉 講師および高知大学 理工学部の 山﨑 朋人 助教と、地球上の光合成生物が持つ「光防御反応(強すぎる光から身を守る反応)」の仕組みについて共同研究を進めていたところ、植物において花芽の形成時期を制御する遺伝子(CONSTANS)が緑藻の光防御反応を制御することを発見しました。詳しい解析の結果、緑藻が強すぎる光から身を守る仕組みには植物の花芽形成と共通する部分があることが分かりました。この発見により、植物の花芽形成に関わる遺伝子系の進化、つまり、遥か昔に水生の藻類が花作りのスイッチを制御する仕組みを獲得していたことが見えてきました。本研究成果は、国際学術誌Nature communications誌に2019年9月10日付で掲載されます。

桜の花と緑藻
【研究の背景】
地球上の光合成生物は、強すぎる光から身を守る「光防御」と呼ばれる環境適応能力を持っています。実際に自然界の日中における太陽の光量(強度)は、多くの場合「光合成に利用できる量」を大幅に超えていることが知られています。私たちが日常的に目にする陸上植物を例にとると、強すぎる光を受け取った葉は「光阻害」と呼ばれる傷害を受け、その程度が大きすぎるとやがて葉は白化し、枯れてしまいます。一度枯れた葉は元どおりに回復することはなく、大きな光阻害を引き起こすような強光が長期に及ぶ、あるいは新たな葉の産生速度が間に合わなければ植物は枯死します。また、肉眼では見えないような単細胞藻類にとっては、強すぎる光はより深刻な問題です。なぜならば、単細胞藻類は細胞内に有する光合成器官(葉緑体)が壊れてしまうと細胞死、つまり個体の死に直結するためです。しかし、自然界に目を向けると、真夏の直射日光のもとでも草木や湖沼の藻類は死滅することなく旺盛に繁茂していることがわかります。これは、「過剰量の光エネルギー」を安全に消去する光防御反応を駆使して強光環境に適応しているためです。
このように、多くの光合成生物にとって共通かつ重要な生存戦略である光防御ですが、その全容は解き明かされていません。研究グループは、これまで単細胞緑藻クラミドモナスにおいて、光防御を引き起こすLHCSR3タンパク質に注目してきました(LHCSR3タンパク質の詳細は2013年5月28日のプレスリリース「過剰な光エネルギーを消去する実体,光合成タンパク超複合体を発見」を参照)。そして、光合成とは直接関係ないとされていた光受容体[*1]の一つであるフォトトロピンがLHCSR3タンパク質の合成に決定的な役割を果たしていることを国際共同研究で発見しました(2016年9月9日のプレスリリース「青色光受容体が光合成にブレーキをかけることを発見~青い光が光合成装置を守る~」を参照)。さらに、青色光を受けたフォトトロピンのシグナルはCUL4-DDB1DET1と呼ばれるE3ユビキチンリガーゼの働きを制御することによってLHCSR3遺伝子の発現を調節していることが明らかになりました(2019年1月1日のプレスリリース「藻類が強すぎる光から身を守るしくみをあきらかに~その根幹部分はヒトにもある?~」を参照)。しかし、光防御のきっかけである「光の受容」から「光防御を担う遺伝子の発現」までの細胞内シグナル伝達系については、まだ多くの謎が残されています。特に、もう一つのLHCSR遺伝子であるLHCSR1は主に青色の光ではなく紫外線によって発現しますが、そのシグナル伝達系についてはほとんどわかっていませんでした。
【研究の成果】
研究グループは、「光の受容」から「光防御を担う遺伝子の発現」までの間に、どのように情報が伝達されるのか、その全容解明を目標としてランダムにマーカー遺伝子を挿入した遺伝子組み換え緑藻を多数作成し、LHCSR1発現変異体の選定と原因遺伝子の特定を進めてきました(Tokutsu et al. 2019 Sci. Rep.)。20,000個体を超える遺伝子組み替え緑藻を調べたところ、強すぎる光のもとで死滅する4つの光防御変異体を単離することに成功しました(図1)。この4つの光防御変異体についてより詳しく調べると、光防御を担う遺伝子・タンパク質が十分に発現しておらず、その結果として光防御反応が正しく駆動していないことがわかりました。このことから、変異体では「光の受容」から「光防御を担う遺伝子の発現」までの間を繋ぐシグナル経路に異常が生じたために、強い光のもとで生存できなくなったと予想されました。
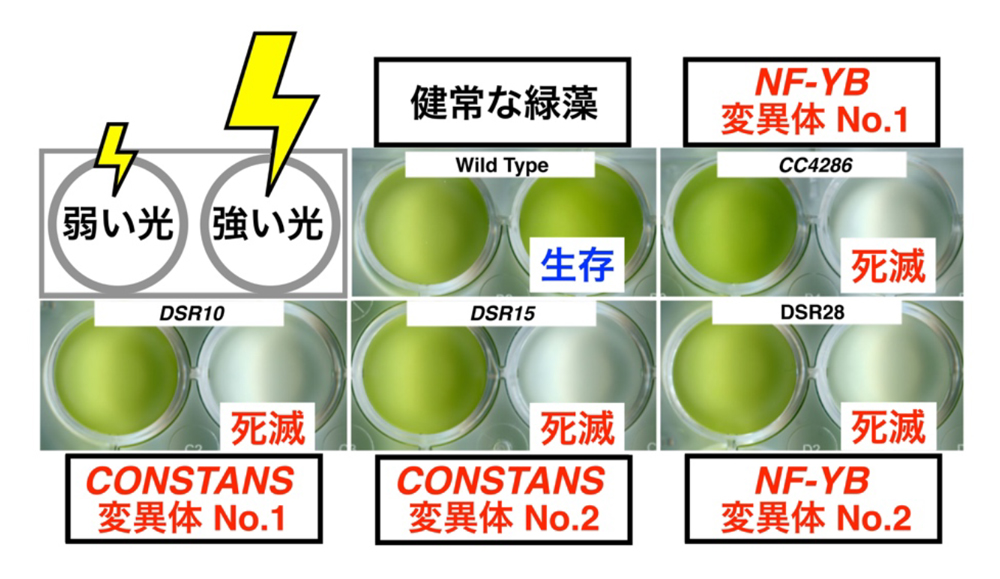
図1 CONSTANSやNF-YBの変異体は強光下で死滅する.
次に、上記の4つの変異体についてゲノム上の変異箇所の特定を試みました。すると驚くことに、2つの変異体ではCONSTANS、残り2つの変異体ではNF-YBと呼ばれる遺伝子が破壊されていることが判明しました。これら2つの遺伝子は、古くから様々な陸上植物において研究されてきた経緯を持ち、花芽の形成を誘導する遺伝子であるFT(FLOWERING LOCUS T)の発現を直接的にコントロールする転写因子であることがわかっています。言うまでもないことですが、水生の微細藻類である緑藻は花を咲かせることはありません。そのため、緑藻におけるCONSTANSやNF-YB遺伝子は、ごく一部の研究者らによってその存在は確認されていましたが、生物学的な役割に関する研究はほとんど行われていませんでした。ところが、今回の実験結果からこれらの遺伝子は緑藻における光防御に必須であることが判明しました。
多くの植物では、その種にとって最適な環境で花を作ることが知られています。これは、その後の開花に伴う受粉率の上昇、つまり、出来るだけ多くの種子を作ることが生存・繁栄に有利であるためと考えられています。従来の研究から、CONSTANSタンパク質は植物に光が当たっている時だけ安定に存在し、FT遺伝子の発現を促すことが知られています。そのため、CONSTANSが季節による日長変化などの環境情報を利用して、植物種ごとに最適な環境での花形成を制御していると考えられています[*2]。では、緑藻はどのようにCONSTANSを利用して光防御をコントロールしているのでしょうか?
研究グループは、光防御変異体の選定過程で得られていた「紫外線の受容体(UVR8)」を失った変異株に着目し、光防御におけるUVR8タンパク質の役割を調べました。その結果、UVR8は紫外線を受け取ると細胞内において核周辺に移動・集積し(図2)、タンパク質の分解を促進するCOP1/SPA1 E3ユビキチンリガーゼ[*3]の活性を抑制することがわかりました。さらに、COP1/SPA1 E3ユビキチンリガーゼの失活はCONSTANSタンパク質の安定化に繋がり、結果として顕著に光防御遺伝子の発現が促進されることを見出しました。
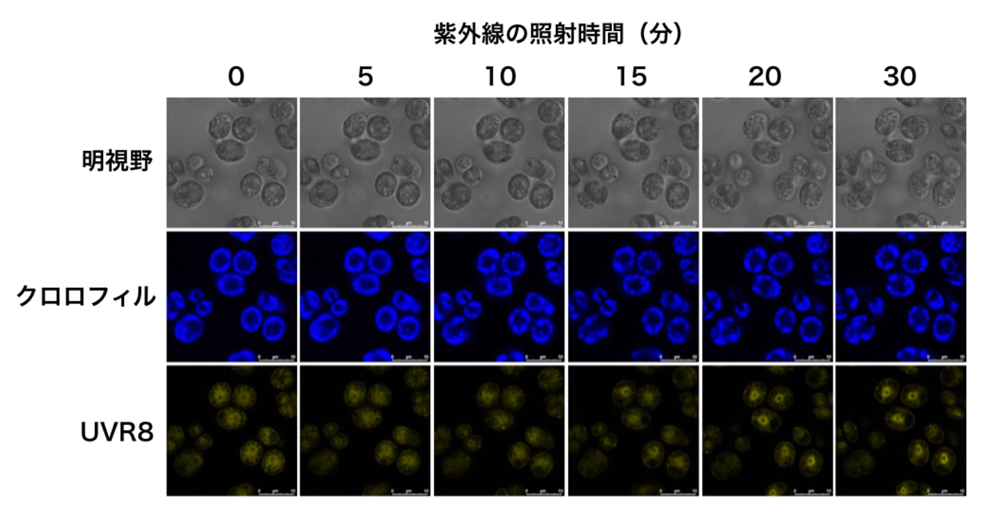
図2 紫外線の照射により核へ集まるUVR8(一番下の黄色いシグナル)
UVR8は核においてCOP1/SPA1 E3ユビキチンリガーゼと相互作用することが分かった.
以上の結果から、これまで謎に包まれていた緑藻の細胞内における「光の受容」から「光防御を担う遺伝子の発現」までの全容が明らかになってきました(図3)。
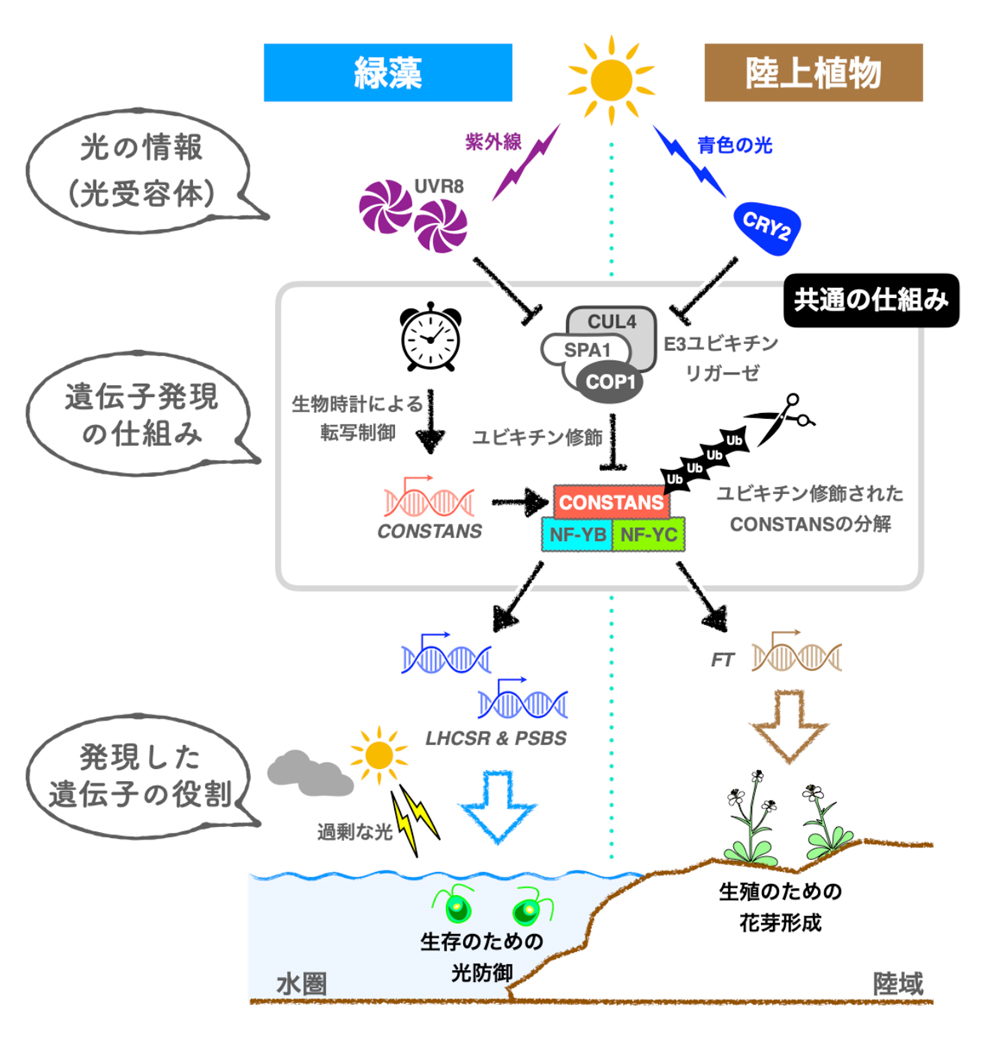
図3 陸上植物の花芽形成と緑藻の光防御との比較
光の受容部と最終的な生物反応を除き、大部分が“共通の仕組み”で制御されている。
図3のように、陸上植物は可視光である青色の光を受け取り、COP1/SPA1 E3ユビキチンリガーゼの活性を抑制する一方、緑藻は紫外線を受け取り、COP1/SPA1 E3ユビキチンリガーゼ活性を抑制します。COP1/SPA1 E3ユビキチンリガーゼ活性の抑制はCONSTANSタンパク質の安定化、そしてCONSTANS/NF-Y転写因子複合体の形成へとつながります。このCONSTANS/NF-Y転写因子複合体は、陸上植物では花作りを始めるためのFT遺伝子の発現、緑藻では強光適応のための光防御遺伝子(LHCSRやPSBS)の発現を制御します。このように本研究では、陸上植物の花作りをコントロールする仕組みが、全く異なる生物反応である緑藻の光防御をコントロールする機能を持つことを発見しました。
【本研究の意義と今後の展望】
本研究で用いられた緑藻クラミドモナスと、現存する陸上植物が属する系統は異なります(図4)。これら2つの系統に属する光合成生物は共通の祖先を持つと考えられていることから、図3で示した「共通の仕組み(COP1/SPA1 E3ユビキチンリガーゼとCONSTANS/NF-Y転写因子を利用した遺伝子発現コントロール)」、つまり、花作りのスイッチの仕組みは、遥か昔に水圏の光合成藻類によって確立されたものと推測できます。

図4 陸上植物と緑藻の進化系統
今日の光合成生物を見ると、花をつける植物や水生の緑藻だけではなくコケやシダを含む多様な種によって構成されており、地球における生物多様性を支えていることがわかります。今後の研究では、本研究で着目した“共通の仕組み”が多様な植物・藻類において、一体どのように「生物種を超えた普遍性」を獲得したのか、そして、地球における生物多様性・進化をどのように運命づけたのか、その謎が解き明かされることが期待されます。
[*1] 光受容体:特定の色(波長)の光を吸収し、光の情報を生体内へ伝えるきっかけを担う分子。植物や緑藻に共通のものとして、青色光受容体フォトトロピンや紫外光受容体UVR8などがある。植物の花芽形成に関与する光受容体としては、青色光受容体の一つであるクリプトクロム(CRY2)が挙げられる。緑藻がCRY2と同じ光受容体を持っているかは未だ明らかにされていない。
[*2] CONSTANSによる日長応答:季節により日照時間(日長)は変化する。そのため、光の受容に伴い安定化される転写因子CONSTANSは、日長が長ければFT遺伝子を発現させる時間が長くなり、結果として植物に花芽形成のためのシグナル物質を多く蓄積させる。花芽の形成には日をまたいで一定程度のFT遺伝子発現が必要であり、その量は植物種によって異なる。そのため、種ごとに花芽をつけるタイミングが異なり、短い日長で花芽を形成するものを短日植物、長い日長で花芽を形成するものを長日植物と呼ぶ。
[*3] E3ユビキチンリガーゼ:特定のタンパク質をユビキチン修飾し、ユビキチンを標的としたプロテアソーム(タンパク質分解酵素)による分解を促進する生体酵素。植物の花芽形成には、COP1・SPA1を内包するE3ユビキチンリガーゼ複合体が深く関与することが報告されている。本研究では、紫外光受容体UVR8がCOP1・SPA1と相互作用することでE3ユビキチンリガーゼ活性が失活する(CONSTANSタンパク質が安定化する)ことが明らかになった。
【掲載誌情報】
雑誌名: Nature Communications(英国9月10日オンライン公開)
論文タイトル: The CONSTANS flowering complex controls the protective response of photosynthesis in the green alga Chlamydomonas
(花成因子CONSTANS 複合体は緑藻クラミドモナスの光合成防御反応を制御する)
著者: 得津隆太郎、鎌田(藤村)このみ、松尾拓哉、山﨑朋人、皆川純
DOI:10.1038/s41467-019-11989-x
【研究グループ】
基礎生物学研究所の得津隆太郎助教、鎌田(藤村)このみ研究員、皆川純教授らのグループ、名古屋大学・遺伝子実験施設の松尾拓哉講師、高知大学・理工学部の山﨑朋人助教からなる共同研究チーム。
【研究サポート】
本研究は、科学研究費補助金である若手研究A、および新学術領域研究「新光合成:光エネルギー変換システムの再最適化」、基礎生物学研究所・生物機能分析室他のサポートを受けて行われました。
【お問い合わせ先】
基礎生物学研究所 環境光生物学研究部門
助教 得津 隆太郎(トクツ リュウタロウ)
教授 皆川 純(ミナガワ ジュン)
【報道担当】
基礎生物学研究所 広報室

