
2023-10-06 東京大学
発表のポイント
- 地球上には30万もの植物種が存在すると見積もられていますが、雌しべと花粉はそれぞれの種に特有のメカニズムによって結びつくため、種の間には「生殖の壁」があると考えられてきました。しかしそのようなメカニズムの多くが謎のままでした。
- 種間の「生殖の壁」をつくる新しい遺伝子Stigmatic Privacy 2(SPRI2)を発見しました。SPRI2は雌しべの核の中で特徴的な粒状構造体を形成しながら細胞壁関連遺伝子を制御する転写因子であり、異種花粉を排除する生殖の壁を作り上げる機能をもつことが明らかになりました。
- SPRI2のような遺伝子を壊して制御すれば、生殖の壁を取り払い新しい作物種を生み出す技術に繋がることが期待されます。

シロイヌナズナ雌しべで発現するSPRI2と、核内において形成されるSPRI2粒状構造体
発表概要
東京大学大学院農学生命科学研究科の藤井壮太准教授(兼任サントリーSunRiSE研究者)と高山誠司教授らによる研究グループは、奈良先端科学技術大学院大学バイオサイエンス領域の伊藤寿朗教授らによる研究グループと共同で、モデル植物のシロイヌナズナから種間の「生殖の壁」をつくるために必要なStigmatic Privacy 2(SPRI2)を同定しました。SPRI2は核内で転写制御因子(注1)として、細胞壁(注2)を形成するために必要な遺伝子を制御して「生殖の壁」を作り出す機能をもつことが明らかになりました。
地球上には多様な植物が繁栄しています。植物種はお互いに影響を与えながら進化してきましたが、なかでも種と種の間で花粉がやり取りされることによって起こる間違った生殖、「繁殖干渉」(注3)が種の存続に大きなマイナスとなることが近年明らかになってきました。そのため、雌しべにおける種間の「生殖の壁」が防御上重要な意義を果たすと考えられますが、そのようなメカニズムはまったく解明されてきませんでした。SPRI2はそのようなメカニズムを制御するマスター因子として本研究で初めて見出されてきました(図1)。雌しべにおけるSPRI2を人為制御することで新しい作物を作り出す技術へと発展することが期待されます。
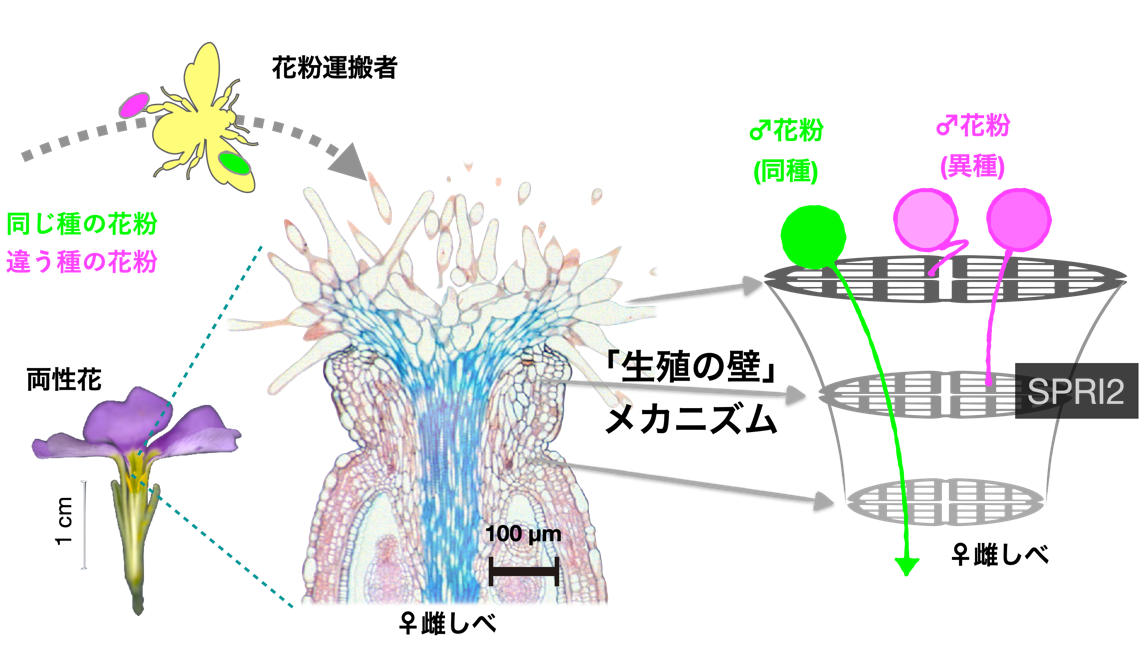
図1:植物の「生殖の壁」とSPRI2について
発表内容
地球上には30万もの植物種が存在すると見積もられています。そのような多様な植物群が存在するにも関わらず、私たち人類はそのうち数百種程度のみを栽培化して農業に利用しています。地球規模の環境問題が迫るなか、可能な限り多くの植物種を遺伝資源として活用してゆくことが望まれます。しかし、植物の雌しべと花粉はそれぞれの種に特有のメカニズムによって結びつくため、種の間には「生殖の壁」が存在します。そのような「生殖の壁」が植物の自由な交雑を妨げ、多様な遺伝資源の活用を難しくしてきました。
その一方で、「生殖の壁」には重要な生物学的意義があることもわかってきました。植物種はお互いに影響を与えながら生態系を形成します。近年、種と種の間で花粉がやり取りされることによって起こる間違った生殖「繁殖干渉」が生じ、種の存続にとって大きなマイナスとなることが段々明らかになってきました。そのため、種間の「生殖の壁」は種を守るために重要な生理機能であると考えられてきました。しかし、そのような「生殖の壁」のメカニズムはほとんど未知であり、応用と基礎両方の観点から解明が望まれていました。
研究グループでは、2019年にそのような「生殖の壁」を形成するStigmatic Privacy 1(SPRI1)という細胞膜タンパク質を発見しました。しかし、SPRI1がどのような機能をもつのか、あるいは「生殖の壁」の形成に関連するその他の因子等は未知でした。そこで、モデル植物のシロイヌナズナを用いたゲノムワイド関連解析(注4)という手法を用いて、異種の花粉を排除するために必要な新しい雌しべ因子の探索を行いました。その結果、第一染色体上に座乗し、異種花粉排除活性と関連性を示すSPRI2を発見しました。SPRI2はSHIと呼ばれる遺伝子転写制御因子ファミリーの一種であり、ゲノム編集法(注5)によってSPRI2の機能を破壊すると、異種の花粉を人工的に授粉した際に花粉管が雌しべに侵入するようになることが明らかとなりました。さらに、SHIファミリーに属するSPRI2と類似のSPRI2-likeタンパク質も異種花粉排除機能をもつことがわかり、SPRI2とSPRI2-likeの二つを同時に破壊することで雌しべにおける種間の「生殖の壁」機能が完全に失われることが見出されました(図2)。従ってこれらの遺伝子群は冗長的に機能しながら「生殖の壁」を制御することが明らかとなりました。また、SPRI2とSPRI2-like以外のSHIファミリー転写因子は異種花粉排除活性には関与せず、これら二つの因子が進化の過程で特有の機能を獲得したことが明らかとなりました。
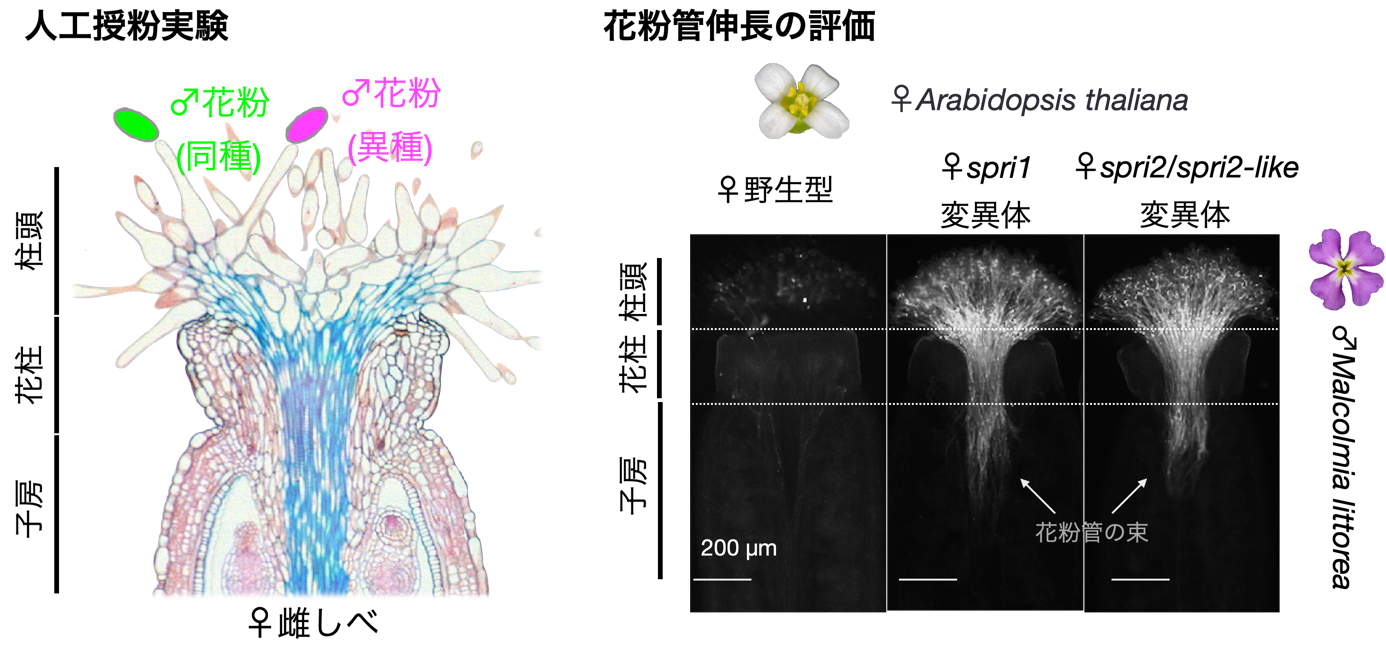
図2:SPRI2を破壊した変異体を用いた人工授粉試験
異種(Malcolmia littorea)を受粉した後の雌しべを固定し、花粉管を蛍光試薬アニリンブルーで染色した。
SPRI2は転写因子であるため、網羅的遺伝子発現解析(注6)を用いてその下流において制御される遺伝子群を探索しました。その結果、SPRI2とSPRI2-likeを同時に破壊した系統では約600個の遺伝子の発現が影響を受け、その多くが細胞壁成分の合成や細胞壁多糖類の化学修飾に関わるものであることが明らかとなりました。さらに、クロマチン免疫沈降解析(注7)によって、TRICHOME BIREFRINGENCE-LIKE 40(TBL40)とTBL45と呼ばれる遺伝子領域のDNAにSPRI2が結合することを見出しました。TBL40/45は細胞壁のペクチンあるいはキシランをアセチル化(注8)する酵素の一種であり、細胞壁の化学修飾が「生殖の壁」の形成に重要な役割を果たす可能性が示唆されました。さらに、SPRI2の変異体ではSPRI1遺伝子の発現が低下する傾向があり、SPRI2が細胞壁修飾酵素やSPRI1など重要な鍵因子の遺伝子発現を司り、「生殖の壁」を作り上げるマスター制御因子であることが示唆されました。
一方、蛍光タンパク質を用いて核内におけるSPRI2の局在を調べたところ、通常のタンパク質とは異なり粒状の構造体として蓄積することが明らかとなりました。光褪色後蛍光回復法(注9)という手法を用いてこの粒状構造に強いレーザーを当てて褪色させたところ、わずか数十秒後に傾向が回復することが明らかとなりました。このことはSPRI2粒状構造体が周辺と分子の交換を行なっており、固体ではなく液体としての性質を有することを示唆しています。また、粒状構造体の形成にはSPRI2タンパク質配列の中のプリオン様ドメインが必須であり、このドメインを欠損させた変異体では異種花粉排除能力が失われることが明らかとなりました。近年、SPRI2粒状構造体のように液―液相分離(注10)と呼ばれる物理現象によって形成される構造体が、生命制御において重要な役割を果たすことが多数報告されています。今回、雌しべにおいてSPRI2粒状構造体が形成されターゲット遺伝子がその場に濃縮されることで、効率的に「生殖の壁」を作り出す「遺伝子発現ファクトリー」が形成されている可能性が示唆されてきました(図3)。
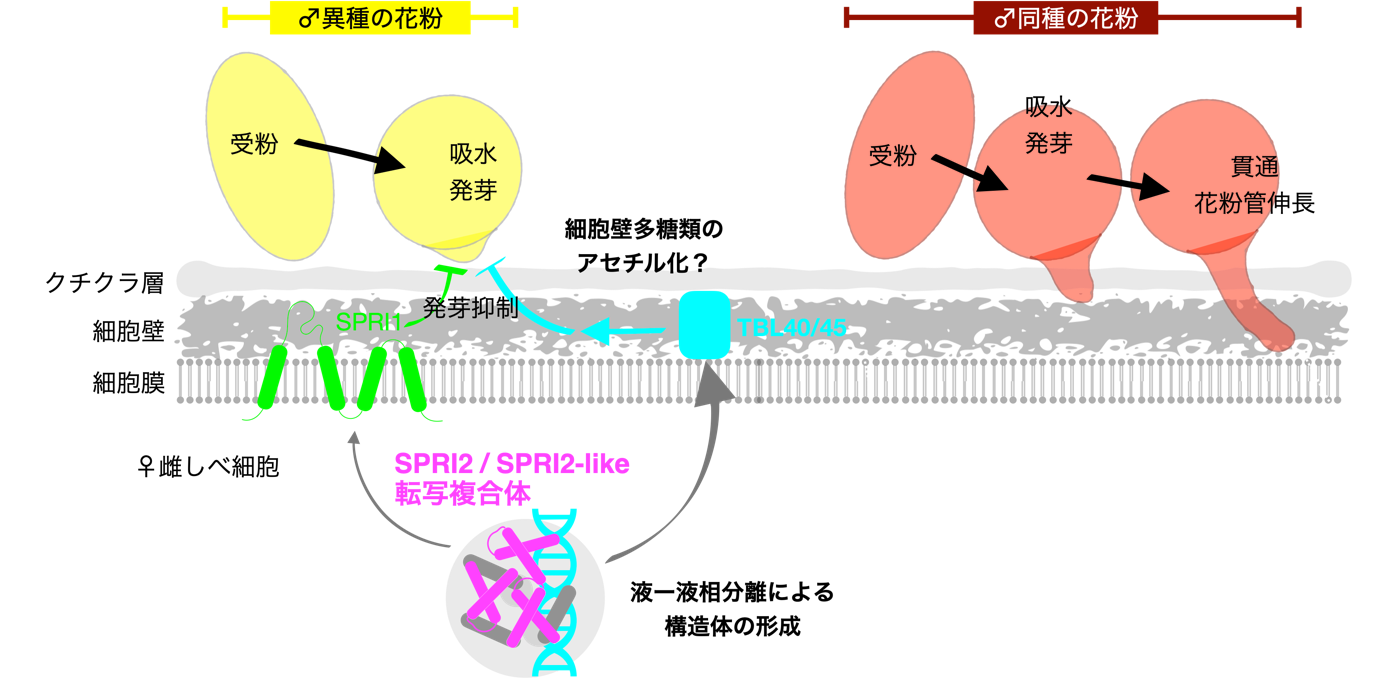
図3:本研究のモデル図
今後は、細胞壁の化学修飾や、SPRI2粒状構造体による転写制御を明らかにすることで「生殖の壁」の更なるメカニズムを解明する必要があります。また、SPRI2のようなマスター因子を作物種において改変することで、人為的に種間ハイブリッドを生み出す技術へと発展することが期待されます。
〈関連のプレスリリース〉
「同種と異種の花粉を区別する分子を発見 ~種の壁を自在に制御する技術の開発に期待~」(2019/07/02)
https://www.a.u-tokyo.ac.jp/topics/topics_20190702-1.html
発表者
東京大学 大学院農学生命科学研究科
藤井 壮太(准教授)<サントリーSunRiSEフェロー>
山本 絵里(修士課程:研究当時)
伊藤 星太郎(修士課程)
タンプラノコン スラチャット(博士研究員:研究当時)
木村 友香(博士研究員)
三浦 大樹(修士課程:研究当時)
加藤 義宣(助教)<JSTさきがけ研究者>
新留 真紀(学術専門職員)
吉田 綾(学術専門職員)
高山 誠司(教授)
奈良先端科学技術大学院大学バイオサイエンス領域
山口 暢俊(准教授)
下里 裕子(博士研究員:研究当時)
和田 七夕子(助教)
伊藤 寿朗(教授)
発表雑誌
- 雑誌
- Nature Plants
- 題名
- SHI family transcription factors regulate an interspecific barrier
- 著者
- Sota Fujii†*, Eri Yamamoto†, Seitaro Ito, Surachat Tangpranomkorn, Yuka Kimura, Hiroki Miura, Nobutoshi Yamaguchi, Yoshinobu Kato, Maki Niidome, Aya Yoshida, Hiroko Shimosato-Asano, Yuko Wada, Toshiro Ito, Seiji Takayama*
†同等貢献、*責任著者 - DOI
- 10.1038/s41477-023-01535-5
- URL
- https://www.nature.com/articles/s41477-023-01535-5
研究助成
本研究は、新学術領域研究・植物新種誕生の原理(課題番号:JP16H06464, JP16H06467)、学術変革領域A・挑戦的両性花原理(課題番号:JP22H05172,JP22H05174,JP22H05176)、新学術領域研究・環境記憶統合(課題番号:JP16H01467, JP18H04776)、学術変革領域A・植物気候フィードバック(課題番号:JP23H04968)、学術変革領域A・不均一環境と植物(課題番号:JP21H05663)、科研費基盤S(課題番号:JP16H06380, JP21H05030)、科研費基盤B(課題番号:JP18H02456)、科研費挑戦的萌芽研究(課題番号:JP15K14626,JP23K17987)、JSTさきがけ(課題番号:JPMJPR16Q8)、サントリーSunRiSE生命科学者支援プログラム等の支援により実施されました。
用語解説
注1 転写制御因子
遺伝子の転写(遺伝情報のRNAへの複写)を調節するための分子のこと。これらの因子はDNA上の特定の領域に結合し、遺伝子の活性化または抑制を制御し、生物の発達、成長、および応答に影響を与える。
注2 細胞壁
植物細胞を包む堅固な外部構造であり、細胞の形状を維持する他に、外部刺激から細胞を保護する役割をもつ。主成分はセルロース、ヘミセルロース、ペクチンなどの多糖類。
注3 繁殖干渉
例えば、ある植物種の雌しべにその近縁種の花粉が付着することで、結実率が低下したりするなど、種と種の間に生じる生殖における相互作用のうち、1種または複数種の繁殖成功度を低下させるもの。
注4 ゲノムワイド関連解析
ある集団において、個体間の形質の違いとDNA配列の違いとの関わりを全ゲノム配列にわたって統計的に検出する手法。
注5 ゲノム編集法
遺伝子やDNA配列を特定の方法で変更する技術の総称。主要な方法にはCRISPR-Cas9などがあり、疾患の治療、新しい特性の追加、農作物の改良など多くの分野で応用される。
注6 網羅的遺伝子発現解析
生物の全ての遺伝子の発現パターンを同時に評価する技術。RNAシークエンシングなどの方法を使用して、特定の条件下でどの遺伝子がどれだけ発現しているかを定量的に測定する。
注7 クロマチン免疫沈降解析
タンパク質とDNAの相互作用を調べる分子生物学的手法の一つ。特定のタンパク質に特異的な抗体を使用して、そのタンパク質がクロマチン内で結合する特定のDNA領域を検出する。
注8 アセチル化
分子構造にアセチル基(CH3CO-)が付加されること。これにより、多糖類の性質や機能が変化し、細胞壁の剛性等に影響を与える。
注9 光褪色後蛍光回復法
生体内の分子や細胞の動態を調査するためのバイオイメージング手法の一つ。特定の蛍光マーカーで標識された分子や細胞を光褪色させ、それから蛍光を再び回復させることで、その拡散速度などを測定する。
注10 液―液相分離
細胞内で特定の生体分子が高濃度で集まり、液体状の微細なドロップレットを形成する現象。これらの液滴内では特定の分子やタンパク質が密集し、特定の生物学的プロセスや反応を効率的に調節する場合がある。
問い合わせ先
〈研究に関する問合せ〉
東京大学大学院農学生命科学研究科 応用生命化学専攻
准教授 藤井 壮太(ふじい そうた)
〈報道に関する問合せ〉
東京大学大学院農学生命科学研究科・農学部
事務部 総務課総務チーム 総務・広報情報担当(広報情報担当)
奈良先端科学技術大学院大学 企画総務課 渉外企画係
