低タンパク質の食事によるエピゲノム変化が遺伝する
2020-03-20 理化学研究所
理化学研究所(理研)開拓研究本部眞貝細胞記憶研究室の吉田圭介協力研究員(研究当時)、石井俊輔研究員らの国際共同研究グループは、マウスを用いて、父親の低タンパク質の食事が生殖細胞でエピゲノム[1]変化を誘導し、精子を通じてそれが子供に伝わり、子供の肝臓における遺伝子発現変化とコレステロールなどの代謝変化を誘導することを明らかにしました。
本研究成果は、「親の食事が子供の成人病などの疾患発症に影響する」という胎児プログラミング仮説[2]のメカニズムを明らかにするもので、生活習慣病などの発症予防につながると期待できます。
胎児プログラミング仮説の現象は、何らかのエピゲノム変化が遺伝することに起因するとされていましたが、そのメカニズムは明らかになっていませんでした。
今回、国際共同研究グループは、野生型雄マウスに低タンパク質食を与えると、その子供の肝臓でコレステロール代謝系遺伝子などの発現が変化するのに対して、転写因子[3]ATF7[4]のヘテロ変異体[5]の雄マウスが父親の場合には、子供に遺伝子発現変化が起こらないことを見いだしました。さらにこのメカニズムとして、低タンパク質食を与えると、雄の精巣の生殖細胞でATF7がリン酸化され標的遺伝子から遊離することで、エピゲノム変化(ヒストンH3K9のジメチル化[6]レベルの低下)が起こり、この変化が精子を経て受精卵に伝わり、遺伝子発現変化を誘導することを明らかにしました。
本研究は、米国の科学雑誌『Molecular Cell』の掲載に先立ち、オンライン版(3月19日付:日本時間3月20日)に掲載されます。

父親の食事が生殖細胞でのエピゲノム変化を誘導し、子供に影響するメカニズム
背景
一連の疫学調査から得られていた「親の世代の栄養状態が子供の疾患、特に糖尿病などの生活習慣病の発症頻度に影響する」という胎児プログラミング仮説が、最近、マウスなどの実験動物を用いて分子生物学的に実証されています。この現象は頻度が高いことから、頻度が低いDNA変異によるものではなく、何らかのエピゲノム変化が生じ、それが遺伝することに起因すると推定されています。しかし、そのメカニズムは明らかになっていませんでした。
石井俊輔研究員らは、これまでに転写因子ATF2の関連因子であるショウジョウバエATF2(dATF2)とマウスなどの動物ATF7が、ストレスなどの環境要因によるエピゲノム変化の誘導に重要な役割を果たすことを明らかにしています注1,2)。そこで、国際共同研究グループは、父親マウスの低タンパク質食が子供に影響する現象におけるATF7の関与について調べました。
注1)2011年6月24日 プレスリリース「親の受けたストレスは、DNA配列の変化を伴わずに子供に遺伝」
注2)2015年9月1日 プレスリリース「自然免疫の記憶メカニズムを解明」
研究手法と成果
国際共同研究グループは、野生型の雄マウスを低タンパク質食あるいはコントロール食で飼育し、コントロール食で飼育した野生型雌マウスと交配し、生まれた子供の肝臓における遺伝子発現パターンを調べました。その結果、雄マウスを低タンパク質食で飼育した場合には、コントロール食で飼育した場合に比べ、421個の遺伝子の発現上昇と101個の遺伝子の発現低下が観察されました(図1左)。発現上昇する遺伝子にはコレステロール代謝系遺伝子などが多く含まれ、父親の低タンパク質食は子供の代謝系に影響することが示されました。
一方、ATF7ヘテロ変異マウスを父親として用いると、ATF7ヘテロ変異体と野生型の子供が生まれますが、いずれの場合も低タンパク質食によって発現が変化する遺伝子はほとんどありませんでした(図1右)。これにより、父親の低タンパク質食が子供に影響する現象にはATF7が必須であることが分かりました。
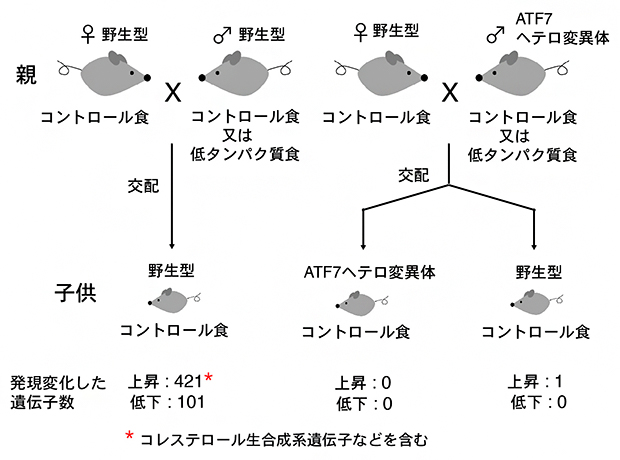
図1 父親マウスの低タンパク質食による子供マウス肝臓での遺伝子発現変化
野生型(左)あるいはATF7ヘテロ変異(右)雄マウスを、低タンパク質食あるいはコントロール食で飼育し、コントロール食で飼育した野生型雌マウスと交配し、生まれた子供をコントロール食で飼育し、その肝臓における遺伝子発現パターンを比較した。野生型雄マウスを低タンパク質食で飼育した場合には、コントロール食で飼育した場合に比べ、多くの発現変化が見られた。ATF7ヘテロ変異雄マウスの場合は、ATF7ヘテロ変異体と野生型の子供が生まれるが、どちらの場合も低タンパク質食によって発現が変化する遺伝子はほとんどなかった。
次に、低タンパク質食がATF7にどのように影響するかを調べたところ、父親の精巣の生殖細胞では、グルタチオン濃度が低下、活性酸素濃度が上昇した結果ストレス応答性リン酸化酵素p38が活性化され、ATF7がリン酸化されることが分かりました。グルタチオンはアミノ酸のシステインを含む小さなペプチドで、細胞内の活性酸素を還元して無毒化する役割を担っています。低タンパク質食はシステイン含有量が少ないため、グルタチオン濃度の低下に伴い活性酸素を還元する能力が低下し、活性酸素濃度が上昇したと考えられます。
また、生殖細胞におけるATF7結合遺伝子を解析した結果、ATF7はゲノム上の約3,500カ所に結合し、多くは遺伝子の転写を制御するプロモーター領域[7]に結合していることが分かりました。そして、低タンパク質食によって80%のATF7結合部位からATF7が遊離し、代表的なATF7結合遺伝子上では、H3K9ジメチル(H3K9me2)レベルの低下(エピゲノム変化)が見られました(図2)。
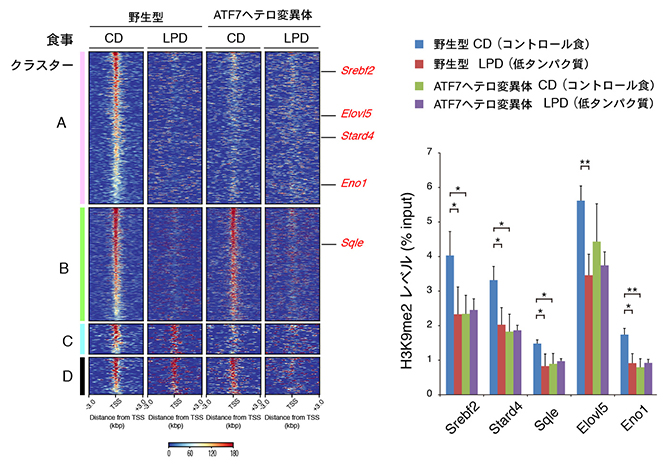
図2 父親マウスの生殖細胞でのATF7結合遺伝子の解析
左:生殖細胞でATF7が結合する遺伝子をChIp-seq法で解析し、転写開始部位を中心にマップした。クラスターAの遺伝子では、野生型低タンパク質食、ATF7ヘテロ変異体のコントロール食・低タンパク質食でATF7が遊離する。クラスターBの遺伝子では、野生型とATF7ヘテロ変異体低タンパク質食でATF7が遊離するが、ATF7ヘテロ変異体コントロール食では遊離しない。
右:生殖細胞での代表的な五つのATF7結合遺伝子上のH3K9me2レベルをqChIP法で調べた。野生型低タンパク質食では、コントロール食に比べてH3K9me2レベルが低下した。
精子が形成される過程では、ほとんどのヒストンは精子固有の核タンパク質プロタミンで置換されますが、完全に成熟した精子にはごく少量のヒストンが残っており、生殖細胞の ATF7結合遺伝子上のヒストンは精子になっても残りやすいことが分かりました(図3左)。また、低タンパク質食によって低下した生殖細胞のATF7結合遺伝子上のH3K9me2レベルは、精子になっても低下したまま保持されることが示されました(図3右)。
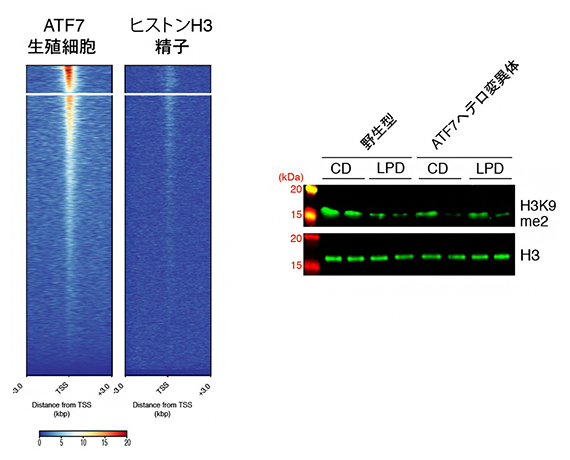
図3 精子でのATF7結合遺伝子の解析
左:生殖細胞におけるATF7結合遺伝子と、精子におけるヒストンH3結合遺伝子をChIP-seq法により解析し、転写開始部位を中心にマップした。両者が相関していることから、生殖細胞のATF7結合遺伝子上のヒストンは、精子になっても残りやすいことが分かった。
右:精子のヒストンH3レベルとH3K9me2レベルをウエスタン法で解析した。野生型低タンパク質食(WTLCD)とATF7ヘテロ変異体コントロール食(CD)・低タンパク質食(LPD)でH3K9me2レベルが低下した。
米国の研究グループの最近の報告で、低タンパク質食により精子中の転移RNA(tRNA)[8]分解フラグメントの量が増加し、それが次世代に伝わり、遺伝子発現を変化させる可能性が示されました注3)。そこで、ATF7のゲノム上の結合部位を調べたところ、生殖細胞ではいくつかのtRNA遺伝子にATF7が結合しており、低タンパク質食により、これらtRNA遺伝子からATF7が遊離し、H3K9me2レベルが低下、tRNA量が増加することが分かりました(図4)。こうして増加したtRNAが精子形成過程で分解され、精子中のフラグメント量が増加すると考えられます。
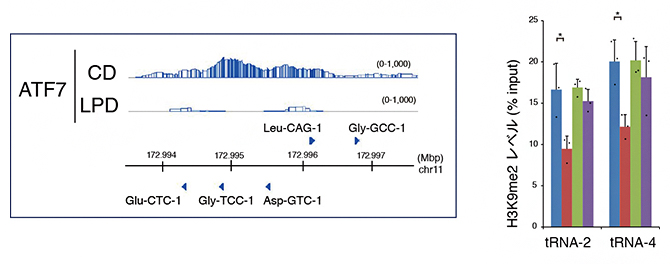
図4 生殖細胞におけるtRNA遺伝子のATF7による制御
左:tRNA遺伝子がクラスターを作るゲノム領域でのATF7結合パターン。低タンパク質食(LPD)によりATF7が遊離する。
右:この領域のH3K9m2レベルをqChIP法で解析した。低タンパク質食により、H3K9m2レベルが低下する。
以上のように、低タンパク質食は雄の生殖細胞において、ATF7結合遺伝子からのATF7の遊離、H3K9me2レベルの低下を誘導し、それが精子を通じて受精卵に伝わり、子供の遺伝子発現を変化させることが示されました。また、ATF7結合遺伝子にはいくつかのtRNA遺伝子も含まれ、低タンパク質食はATF7を介してtRNA量とtRNA分解フラグメント量を増加させ、それが受精卵に伝わることもメカニズムの一端と考えられます。
注3)Sharma U. et.al. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science. 2016 Jan 22;351(6271):391-396.
今後の期待
本研究により、「親の食事が子供の成人病などの疾患発症に影響する」という胎児プログラミング仮説のメカニズムの一端が解明されたことにより、どのような栄養条件が糖尿病など子供の生活習慣病の発症に影響するかを科学的に解明する手掛かりが得られました。本成果は今後、より健康的な栄養条件の解明やサプリメントなどの開発につながると考えられます。
また、生殖細胞でエピゲノム変化が誘導される遺伝子群が同定されたことにより、これらの遺伝子のエピゲノム変化を測定することで、次世代での代謝変化を予測することも可能になると期待できます。
補足説明
1.エピゲノム
接頭辞「エピ(付加したの意)」と「ゲノム」をつないだ言葉で、DNAやDNAが巻き付くヒストンタンパク質がメチル化など化学修飾された遺伝子配列情報のこと。エピゲノムのいくつかは細胞分裂を越えて伝わる。また、エピゲノムは環境要因により変化し、それがさまざまな疾患の発症に影響することが示唆されている。
2.胎児プログラミング仮説
「低出生体重児は、成人期に糖尿病などの生活習慣病発症リスクが高い」という疫学調査の結果をもとにした「将来の健康や疾患発症リスクは、胎児期や生後早期の環境の影響を受けて決定される」という仮説。食物やストレスなど後天的な要因によって起こるエピゲノム変化が、疾患発症のリスクに影響すると考えられている。そしてこのエピゲノム変化は世代を越えて遺伝しうることが議論になっている。
3.転写因子
DNAに配列特異的に結合するタンパク質で、プロモーターやエンハンサーといった転写制御領域に結合し、RNAポリメラーゼによる遺伝子の転写を活性化あるいは不活性化する。
4.ATF7
ATF2と構造が類似した転写因子。ATF2は30年前に石井俊輔研究員らのグループにより初めて同定された。ATF2やATF7は多様なストレスに呼応して、ストレス応答性リン酸化酵素p38でリン酸化されるという特徴を持つ。
5.ヘテロ変異体
二つのゲノムのうち、一つだけが変異を持つ個体。したがって、ATF7ヘテロ変異体でのATF7の量は野生型の半分になっている。ヘテロ変異体雄マウスを野生型雌と交配させると、ヘテロ変異体と野生型の子供が生まれる。
6.ヒストンH3K9のジメチル化
ヒストンH3のN末端から9番目のリジン(K)のジメチル化(H3K9me2)のこと。H3K9me2は転写が不活発で、固い安定なクロマチン構造を形成する。このような構造は、体細胞分裂や生殖細胞での減数分裂を経ても安定に維持される。
7.プロモーター領域
ゲノムDNA上でRNAに書き写される領域の一番上流にあり、遺伝子を発現させる機能を持つ部分をプロモーター領域(配列)という。
8.転移RNA(tRNA)
転移リボ核酸(transfer RNA)の略号。アミノ酸の種類ごとに1~数種類のtRNAが存在する。tRNAはそれぞれ対応するアミノ酸を末端に結合し、リボソームまで運搬する。リボソームにアミノ酸を受け渡すと、再びアミノ酸を結合して、リボソームまで運搬する働きを繰り返す。
国際共同研究グループ
理化学研究所 開拓研究本部 眞貝細胞記憶研究室
研究員 石井 俊輔(いしい しゅんすけ)
協力研究員(研究当時) 吉田 圭介(よしだ けいすけ)
研究員(研究当時) 前川 利男(まえかわ としお)
研修生(研究当時) ニュン・ホン・リー(Nhung Hong Ly)
筑波大学医学医療系
准教授 村谷 匡史(むらたに まさふみ)
大学院生 藤田 晋一郎(ふじた しんいちろう)
大阪大学 蛋白質研究所
教授 岡田 眞里子(おかだ まりこ)
大学院生 安藤 美波(あんどう みなみ)
九州大学大学院 医学研究院
教授 伊藤 隆司(いとう たかし)
准教授 三浦 史仁(みうら ふみひと)
研究員 荒木 啓充(あらき ひろみつ)
東京大学 定量生命科学研究所
教授 白鬚 克彦(しらひげ かつひこ)
助教 加藤 由紀(かとう ゆき)
ストラスブール大学
教授 ブルーノ・シャットン(Bruno Chatton)
研究支援
本研究は、科学技術振興機構(JST)戦略的創造研究推進事業CREST「エピゲノム研究に基づく診断・治療へ向けた新技術の創出(研究総括:山本雅之)」の研究課題「環境要因によるエピゲノム変化と疾患(研究代表者:石井俊輔)」による支援を受けて行われました。
原論文情報
Yoshida K, Maekawa T, Ly NH, Fujita S, Muratani M, Ando M, Katou Y, Araki H, Miura F, Shirahige K, Okada M, Ito T, Chatton B, and Ishii S., “ATF7-dependent epigenetic change is required for intergenerational effect of paternal low-protein diet.”, Molecular Cell, 10.1016/j.molcel.2020.02.028
発表者
理化学研究所
開拓研究本部 眞貝細胞記憶研究室
協力研究員(研究当時) 吉田 圭介(よしだ けいすけ)
研究員 石井 俊輔(いしい しゅんすけ)
報道担当
理化学研究所 広報室 報道担当

