2021-03-12 理化学研究所
理化学研究所(理研)生命機能科学研究センター形態形成シグナル研究チームのチュ・ウェイチェン研究員と林茂生チームリーダーの研究チームは、ショウジョウバエの飛翔器官が構築される過程で筋肉と骨格を機能的に連結させる仕組みを研究し、両者を連結させる腱組織の構築において、細胞外基質[1]が強靱な繊維に変換されることを解明しました。
腱組織の発達は筋肉からの力学的刺激により促進されることが幅広い動物種で知られており、本研究成果は、ヒトの発達期における筋損傷などで起こる運動器官の発達不全の発症機構の理解に向けた基礎研究に貢献すると期待できます。
昆虫の飛翔を駆動する強力な飛翔筋[2]は、腱組織を介して外骨格(クチクラ[1])に接続し、骨格を動かすことで翅のはばたき運動を駆動します。飛翔筋は発生途中においても強大な収縮力を発揮するため、腱組織が相応の強度を獲得する必要があります。しかし、腱を構築する細胞外基質がどのようにして強度を獲得するのかは不明でした。
今回、研究チームは、腱組織の細胞外基質のタンパク質Dumpy[3]が筋肉からの張力に誘導されて、強靱な繊維に転換することを発見しました。さらに、もう一つの細胞外基質のタンパク質Quasimodo(Qsm)[3]がDumpyの強度を増強することを見いだしました。
本研究は、科学雑誌『Current Biologyオンライン版(2月4日付)に掲載されました。

正常型ショウジョウバエ(左)と腱と翅の形成が正常に進まないdumpy, qsm二重変異体
背景
筋肉の収縮によって生み出された力は、腱を通して骨に伝えられます。この筋・腱・骨による運動の仕組みは、内骨格を持つ脊椎動物と、外骨格を持つ節足動物で共通です。哺乳類の胎児は、発生中の運動器官を動かすことで筋肉と骨格の機能を高め、生後直ちに運動できるように準備します。つまり赤ちゃんの胎動は、お母さんの子宮の中での準備体操なのです。しかし、脆弱な胎児の体で筋肉だけが一気に大きな力を発揮すれば、その力を受け止める骨格とそれらをつなぐ腱が壊れてしまいます。したがって、筋肉の力の増大に対応して、骨格と腱が協調して強度を高める必要があります。
脊椎動物では、運動の刺激に応答して骨芽細胞の活性が上昇することで骨の形成が促されることがよく知られています。一方、腱の発生においても、機械的刺激が損なわれた運動器官では腱の発達が遅れ、先天的な運動機能異常の原因となることが分かっています。腱の本体は、コラーゲン[1]などの繊維化した細胞外基質タンパク質が筋肉と骨格をつなぐ、強靱な細胞外構造です。しかし、細胞の外にある腱構成タンパク質の繊維化が、どのようにして機械刺激に応答するのかはよく分かっていませんでした。
そこで研究チームは、節足動物であるキイロショウジョウバエの蛹で発達する飛翔筋システムを腱形成のモデルとし、腱構造を構築する繊維状のタンパク質が形成される仕組みの解明に取り組みました。
研究手法と成果
昆虫の腱構造は、特殊化した外骨格が筋肉と結合したものです。ハエの飛行は、強力な間接飛翔筋[2]が胸郭の外骨格を高周期で動かすことで、胸郭に接続された翅を駆動させます(図1)。
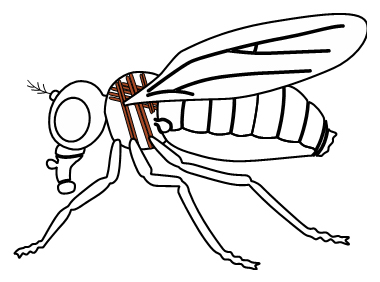
図1 ショウジョウバエの間接飛翔筋
ショウジョウバエ成虫の間接飛翔筋(茶色)の模式図。筋の両端が胸郭の外骨格に結合し、収縮により外骨格を変形させることで翅を動かす。間接飛翔筋には、体の前後方向に収縮するDorsal longitudinal muscle(DLM)と、背腹方向に収縮するDorsal ventral muscle(DVM)の二つがあり、本研究ではDLMの腱構造を観察した。
研究チームは細胞外基質のタンパク質Dumpyに着目し、蛹期において発生中の間接飛翔筋に接続する腱構造の発達を観察しました(図2左)。蛹中で発生する成虫の間接飛翔筋は、表皮の一部である腱細胞に体内から接続し、腱細胞は表面側で蛹のクチクラに固定されています。蛹発生24時間頃になると、脱皮の前段階として表皮/腱細胞と蛹のクチクラの間に「間隙」が生じます。その後12時間のうちに、発生中の間接飛翔筋からの収縮力により間隙は拡大します(図2中)。緑色蛍光タンパク質(GFP)で標識されたDumpyをライブイメージング[4]で観察すると、間隙の中にまず網目状に蓄積し、間隙の拡大に伴って、筋による引っ張り方向に沿った繊維状に変化することが観察されました(図2右)。また、Dumpyの繊維化は発生中の間接飛翔筋からの収縮力に依存して進行することが分かりました。
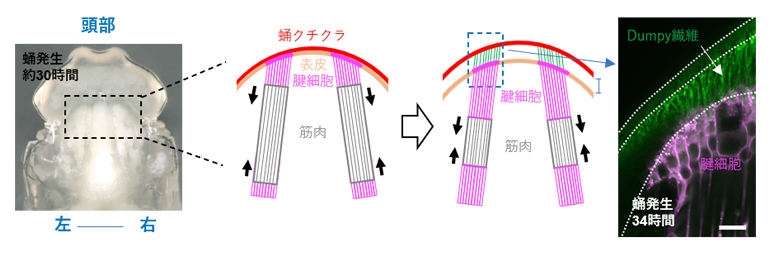
図2 間接飛翔筋に接続する腱構造の発達
左)蛹発生約30時間の頭胸部を背側から見た図。間接飛翔筋の前部の結合部分を四角で囲んだ。
中)左図の四角で囲んだ領域で発達中の腱構造の模式図。表皮の一部である腱細胞は、初めは蛹クチクラに固定されている。その後、脱皮の前段階として蛹クチクラと表皮の間に間隙が生じると、Dumpyタンパク質が間隙に蓄積する。
右)間隙中に蓄積するDumpyタンパク質。筋による引っ張り方向に沿った繊維状の構造を示した。スケールバーは10マイクロメートル(μm、1μmは1,000分の1mm)。
Dumpyの機能が低下するdumpy変異体では、胸郭などの形態に異常が見られます。研究チームは、これに類似した変異表現型を示すquasimodo(qsm)変異体に着目しました。dumpyとqsmの二重変異体を作製すると、それぞれの変異表現型が強調されることから、両遺伝子は協調して働くことが明らかになりました。そこで、qsm変異体においてDumpyの繊維化をライブイメージングにより観察したところ、筋肉からの張力が増すにつれてDumpy繊維が過剰に伸長し、最大張力の時点において切断する現象が観察されました(図3)。これにより、腱上皮は筋肉の力で陥没し、その状態で成熟するために羽化時の陥没した胸郭の表現型を成すことが分かりました(1ページ目の写真右)。
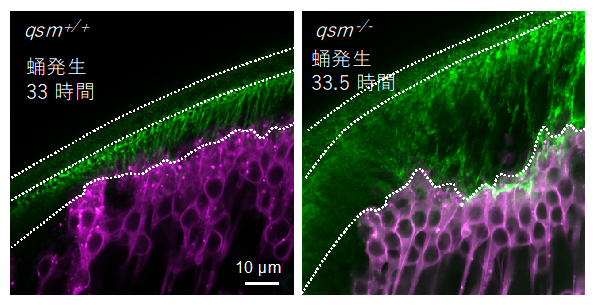
図3 qsm変異体におけるDumpy線維の異常
Dumpy線維(緑)と腱細胞(マゼンタ)の観察像。左側は正常型のショウジョウバエ。右側のqsm変異体では、間隙中のDumpy繊維が過剰に伸長・切断し、腱上皮が陥没した(不規則な波線で示す)。
DumpyとQsmはどちらも、タンパク質重合作用を示すZPドメイン[3]と呼ばれる構造を持つ細胞外基質タンパク質です。しかし、Dumpyは最大22,949個のアミノ酸から構成される巨大タンパク質であるのに対して、Qsmは414個のアミノ酸からなる標準的なサイズです。タンパク質の動態を蛍光マーカーを用いて観察したところ、両者は腱細胞で生産され、細胞内小胞で相乗りした形で輸送されることが分かりました。細胞外に放出されたDumpyは不溶性の繊維となる一方で、Qsmは組織外を広く拡散する様子が観察されました(図4)。培養細胞を用いた実験により、DumpyのZPドメインとQsmは細胞内では安定して結合するものの、細胞外に放出された後には解離することが明らかになりました。
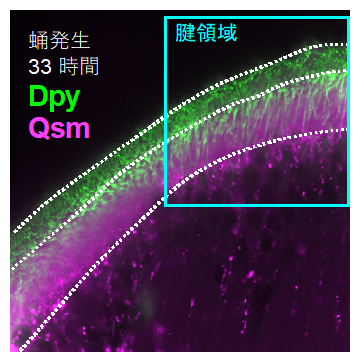
図4 DumpyとQsmの動態
Dumpyタンパク質(緑)とQsmタンパク質(マゼンタ)の観察像。DumpyとQsmは異なる動態を示すが、腱領域などで局在が重なる。またQsmは組織内を広く拡散している。
細胞外を拡散するQsmがどのように機能するかを調べるために、qsm変異体においてQsmを局所的に発現させました。すると驚いたことに、胸郭部の表皮細胞や筋肉で発現させたQsmは、翅の形成異常を含む全てのqsm変異表現型を正常に戻すことができました。これは、分泌されたQsmが組織の間隙を拡散し、遠く離れた標的器官でもDumpyの繊維化を促進したためと考えられました。実際に、筋肉で発現させた蛍光標識Qsmは体腔の中を拡散し、腱細胞に取り込まれた後にDumpyを含む細胞内小胞に局在することが確認されました。
成熟したDumpy繊維は、タンパク質が周期的に配列した繰り返し構造をとることが見いだされ、タンパク質配列の規則化が強靱な繊維構造を生み出すものと考えられました(図5)。さらに、Dumpyの持つZPドメインは自律的に重合化して、高次構造をとることが分かりました。しかし、QsmのZPドメインには重合化活性がなく、細胞内に限ってDumpyのZPドメインと相互作用できることが分かりました。これは、Qsmは細胞外を拡散し、標的細胞に取り込まれることで、Dumpyの成熟を促進させる機能を果たすことによると考えられ、細胞外基質タンパク質の新たな機能として注目されます。
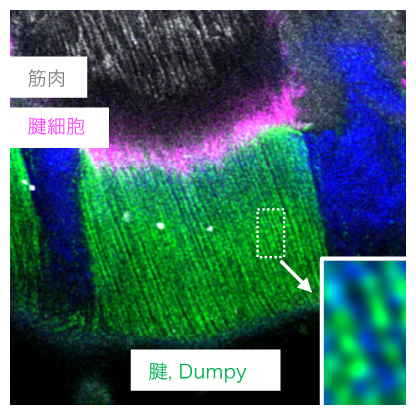
図5 Dumpy繊維の周期的配列構造
Dumpyタンパク質は幼虫にも発現する。Dumpy繊維(緑)が幼虫のクチクラ(青)に入り込む様子を示す。十分に線維化した領域(四角)では、Dumpyの蛍光シグナルがスポット状に観察され、タンパク質が周期的に配列した繰り返し構造をとっていることが分かった。
今後の期待
本研究から、細胞外基質タンパク質Dumpyが、筋肉からの引っ張りによる機械的作用と、Qsmタンパク質との生化学的な作用によって強靱な繊維として成熟することで、強力な飛翔筋による腱組織の変形を回避していることが明らかとなりました。
細胞の外で生体高分子が、どのようにして集合して機能的な高次構造を構築するのかは未解明の課題です。本研究はその過程に関わる物理的要素(引張り力)と化学的要素(Qsmの作用)を見いだしたことで、細胞外マトリックス研究に新たな展望を開きました。さらに、可溶性のQsmがDumpy繊維の成熟を促進させる知見は、細胞外マトリックスの形成を操作する手法につながる可能性を示しています。
腱組織の発達は、筋肉からの力学的刺激により促進されることが幅広い動物種で知られており、今回得られた知見は、ヒトの発達期における筋損傷などで起こる運動器官の発達不全の発症機構の理解に向けた基礎研究に貢献すると期待できます。
補足説明
1.細胞外基質、クチクラ、コラーゲン
細胞外基質は細胞と細胞の間を満たし、生体組織を包み込む高分子の構造体。組織の支持体となるだけでなく、細胞外環境の情報を伝えることで細胞の増殖や分化を制御する。コラーゲンは代表的な細胞外基質の一つで、脊椎動物では真皮や腱などに見られる。昆虫の体表面を覆うクチクラは、表皮細胞が分泌した細胞外基質で、丈夫な外骨格を形成する。
2.飛翔筋、間接飛翔筋
飛翔筋は昆虫の翅を駆動する筋肉のこと。飛翔筋には、翅の基部に接続し翅を動かす直接飛翔筋と、外骨格に接続し間接的に翅を動かす間接飛翔筋の2種類がある。
3.Dumpy、Quasimodo(Qsm)、ZDドメイン
Dumpyタンパク質をコードするdumpyは、ショウジョウバエで発見された遺伝子で、変異体の翅が野生型に比べてずんぐりとした形状をとることから命名された。ショウジョウバエゲノム最大の遺伝子で、タンパク質の分子量は250万を超える。Quasimodoタンパク質をコードするquasimodoは、概日リズムの光依存的同調に関わる膜タンパク質の遺伝子として同定された。両者が共通して持つZPドメインは、哺乳類卵を取り囲む膜(透明帯:zona pellucida)を構成する細胞外基質に共通する約260個のアミノ酸配列である。
4.ライブイメージング
生きた細胞や組織のさまざまな生命活動を継時観察すること。特に、GFPなどの蛍光マーカーを用いて特定の分子や細胞、組織を標識し、蛍光顕微鏡でその動きや変化を詳細に観察する技術。
研究支援
本研究は、台湾科技部補助赴國外從事博士後研究「Study the role of zona pellucida domain(ZPD) proteins in apical extracellular matrix during epithelial morphogenesis and wound healing(チュ・ウェイチェン)」および日本学術振興会(JSPS)科学研究費補助金挑戦的研究(開拓)「細胞外基質ナノ構造の生物学的構築原理の解明(研究代表者:林茂生)」による支援を受けて行われました。
原論文情報
Wei-Chen Chu, Shigeo Hayashi, “Mechano-chemical enforcement of tendon apical ECM into nano-filaments during Drosophila flight muscle development”, Wei-Chen Chu, Shigeo Hayashi, 10.1016/j.cub.2021.01.010
発表者
理化学研究所
生命機能科学研究センター 形態形成シグナル研究チーム
研究員 チュ・ウェイチェン(Chu Wei-Chen)
チームリーダー 林 茂生(はやし しげお)
報道担当
理化学研究所 広報室 報道担当

