
2022-02-09 理化学研究所,大阪大学,東京大学,東京農業大学
理化学研究所(理研)環境資源科学研究センター機能開発研究グループのキム・ジュンシク研究員(研究当時、現バイオ生産情報研究チーム研究員)、篠崎一雄グループディレクター、大阪大学大学院理学研究科生物科学専攻の坂本勇貴助教、東京大学大学院新領域創成科学研究科先端生命科学専攻の松永幸大教授、東京農業大学農生命科学研究所の篠崎和子教授らの共同研究グループは、環境変動に応じた植物の根の伸長調節に関わる新たな遺伝子制御因子を発見しました。
本研究成果は、植物工場や都市型農業に向けた地下部生産性の向上に貢献すると期待できます。
今回共同研究グループは、「小胞体ストレス応答(UPR)[1]」と呼ばれる細胞内恒常性の維持機構が欠損したシロイヌナズナの変異株(bz1728)が示す、著しい根の伸長阻害を回復した突然変異体nobiro6株の分子遺伝学的解析を行いました。その結果、伸長回復は基本転写因子複合体[2]の構成因子「TAF12b」の機能欠損によることが明らかになりました。さらに、網羅的遺伝子発現解析[3]を行ったところ、bz1728株では数百個の遺伝子の発現が上昇・低下の両方向に大きく変動するのに対し、nobiro6株では上昇方向の変動だけが通常程度に回復することが分かりました。回復した遺伝子群の多くがストレス耐性獲得に機能することから、TAF12bは植物が感知した外部ストレスのシグナルを根の細胞の成長応答に結び付ける重要な遺伝子制御因子であると考えられます。
本研究は、科学雑誌『Proceedings of the National Academy of Sciences of the United States of America』(2月8日付)への掲載に先立ち、オンライン公開されました。
背景
地中に根を張る陸上植物は最適な環境に自ら移動できないため、高温や乾燥、病害などさまざまな環境ストレスに対処する複雑な応答機構を発達させてきました。このストレス応答機構では、環境ストレスへの耐性が高まる反面、植物体の成長が抑えられることが分かっています。しかし、環境ストレスに対する特定の組織における成長抑制の分子メカニズムは明らかになっていませんでした。
共同研究グループは、細胞内ストレス応答機構の一つである「小胞体ストレス応答(Unfolded Protein Response:UPR)」の活性化による、植物の根の伸長成長の抑制現象を分子遺伝学的に解明する研究を進めてきました。UPRの活性化は、生物内外の環境悪化によって細胞内に蓄積した不完全タンパク質を感知することで起こります。UPRには、真核生物共通の細胞内恒常性を維持する役割とともに、外部ストレスを細胞内シグナルに変換して遺伝子発現制御に伝える細胞内ストレスセンサーとしての役割があると考えられています。
分子遺伝学のモデル種であるシロイヌナズナ(Arabidopsis thaliana)では、三つの転写因子[4](bZIP17、bZIP28、bZIP60)がUPRの制御に関わっています。2018年にキム・ジュンシク研究員らは、そのうちのbZIP17とbZIP28を同時に機能欠損させた変異株(bz1728)では、根の伸長が野生株に比べて10%程度に阻害されることを見いだしました注)。しかし、その表現型は根の伸長阻害を特徴とする他の変異株と関連性が乏しく、また植物ホルモンなど根の伸長成長に重要な代謝経路においても特異性のある変化は見られませんでした。これらのことから、bz1728株が示す根の伸長阻害は新しい遺伝因子に起因すると考えられ、本研究を行いました。
注)2018年2月6日プレスリリース「植物の根の伸長を支えるストレス応答機構」
研究手法と成果
共同研究グループは、bz1728株における根の伸長阻害に関与する新たな遺伝因子を探索するため、ゲノム上にランダムな突然変異を誘導した突然変異集団を作出しました。親株であるbz1728から根の伸長成長を回復した一連の変異株を選抜し、「nobiro(ノビロー)」と名付けました(図1)。

図1 根の伸長が阻害されたbz1728株とそれを回復したnobiro6株、野生株の表現型
転写因子bZIP17とbZIP28を同時に機能欠損させた変異株bz1728(中央)では著しく根の伸長が阻害されるが、bz1728株の変異株の一つnobiro6(右)は、根の伸長成長が回復している。
本研究では、nobiroの一つであるnobiro6株を対象に原因遺伝子を特定し、根の伸長回復の分子メカニズムを解明するための分子遺伝学解析を行いました。
まず、nobiro6株とbz1728株間の戻し交配[5]から得られた後代(子孫)を、さらに自殖して得られたBC1F2[5]分離集団を対象にゲノム再解読[6]を行いました。根の伸長度合いが異なる複数の個体から解読した遺伝子変異データを比較解析したところ、根の伸長回復と相関して検出される突然変異の一つが、基本転写因子複合体の構成因子の一つである「TAF12b」の正常な発現を阻害することを発見しました。さらにゲノム編集法[7]を利用してbzip17、bzip28、taf12b遺伝子の多重の機能欠損変異株を作出したところ、nobiro6株と同程度の根の伸長回復を示したことから、このtaf12b遺伝子上の変異がnobiro6株の伸長回復の原因であることが証明されました。
TAF12bが遺伝子の発現量を調節する転写因子の一種であることから、bz1728株からnobiro6株への遺伝子発現量の変化が根の伸長回復をもたらしたと予想できます。そこで、野生株とbz1728株、nobiro6株を比較するための網羅的遺伝子発現解析を行ったところ、bz1728株では、野生株に比べて数百個の遺伝子の発現が上昇・低下の両方向に数百倍にいたる大きな変動を示しました(図2左)。一方、nobiro6株では上昇方向の発現変動が野生株と同程度まで回復しているのに対して、低下方向の発現変動はbz1728株の水準のまま保持されていることが分かりました(図2右)。このことから、bz1728株が示す上昇方向の遺伝子発現変動に、根の伸長成長を抑制する機構が含まれている可能性が示されました。
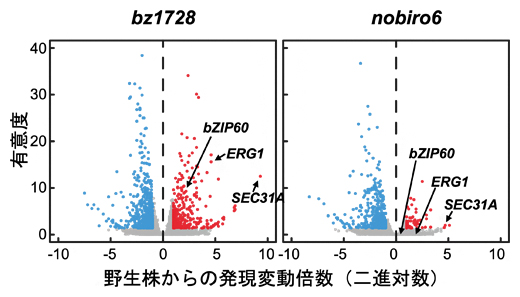
図2 bz1728株およびnobiro6株における網羅的遺伝子発現変動
各点は、野生株に比べて一つの遺伝子の発現量がどれくらいの有意度(縦軸)を持って、どの程度(横軸)変動したかを表す。赤は野生株に比べて発現量が有意に上昇した遺伝子を、青は有意に低下した遺伝子を表す。矢印はbZIP60遺伝子とその代表的なターゲット遺伝子(ERG1、SEC31A)の発現量を示す。
シロイヌナズナのUPR上で機能する転写因子には、bZIP17とbZIP28のほかにbZIP60があります。bz1728株ではbZIP60遺伝子とその下流遺伝子の発現が過剰に活性化することから、bz1728株が示す上昇方向の遺伝子発現変動はbZIP60とTAF12bの共活性によると予想しました。そこで、細胞実験系を通して、bZIP60とTAF12bが物理的に相互作用すること、bZIP60の転写活性化能がTAF12bの添加によりさらに強化されることを証明しました。
さらに、TAF12bの単一機能欠損株(taf12b株)の生理学および分子遺伝学的特性を調べました。すると、taf12b株では、人為的に誘導された小胞体ストレスによる根の伸長抑制応答が鈍くなっており、UPRの活性も低下していることが分かりました。
以上のことから、nobiro6株の原因変異が見つかったTAF12bが、植物の小胞体ストレス応答における転写補助因子[8]としてストレス信号に応じたUPRの活性を助長し、ストレス応答性の根の伸長抑制に機能することが明らかになりました。
今後の期待
今後、他のnobiro株に対する分子遺伝学研究を進めることで、UPRによる根の伸長制御機構に対する理解がさらに深まると考えられます。これらの研究成果は、環境と植物の相互作用に対する学問的価値にとどまらず、ダイコンやサツマイモなど、根を利用する作物の改良を通して、植物工場や都市型農業に向けた作物の生産性向上に貢献できる可能性があります。
従って今回の研究は、国際連合が2016年に定めた17項目の「持続可能な開発目標(SDGs)[9]」のうち「2.飢餓をゼロに」や「13.気候変動に具体的な対策を」などへの貢献が期待できます。
補足説明
1.小胞体ストレス応答(UPR)
小胞体は真核生物の細胞内に存在する細胞小器官の一種。生命現象の維持に必須なタンパク質の加工(折り畳み)や糖鎖などの修飾が行われ、細胞の工場に例えられる。タンパク質の加工プロセスは外部環境の変化にとても敏感で、高温や乾燥、病害などのさまざまな刺激を受けると、正しく加工されていない不完全タンパク質を生産する。小胞体は不完全タンパク質の過剰な蓄積をストレスの一種と認識し、その程度に応じてタンパク質加工プロセスを強化する遺伝子制御機構を装備しており、これを小胞体ストレス応答と呼ぶ。UPRはUnfolded Protein Responseの略。
2.基本転写因子複合体
ゲノム上の遺伝子領域から、タンパク質となる遺伝情報をメッセンジャーRNAとして読み取る過程を転写という。基本転写因子複合体は、転写を行うRNAポリメラーゼⅡと相互作用しながら転写を開始する一連の転写関連タンパク質の複合体である。TAF12は、これらの複合体のうち最も巨大なTFIIDの構成因子の一つ。
3.網羅的遺伝子発現解析
一つの細胞には数万から数十万の遺伝子が存在する。これらの全遺伝子の発現を網羅的に解析するためのさまざまな方法が開発されている。近年では、比較的短い(100~300塩基)配列断片を大量(100億塩基以上)に生産する次世代シーケンス技術の利用が主流になっている。
4.転写因子
一つの細胞に存在する数万の遺伝子は、適材適所で機能するためにその発現が制御されなければならない。転写因子はタンパク質の一種で、DNAと直接相互作用することで標的遺伝子の発現を抑制、または誘導する。各転写因子は固有のターゲット塩基配列を持っていて、細胞内外の状況に応じて適切な遺伝子の発現制御に寄与する。
5.戻し交配、BC1F2
子世代を親世代のどちらかと交雑することを戻し交配(backcross:BC)と呼ぶ。1回のBCにより得られた雑種第一代(the first filial:F1)を自殖して得られた後代がBC1F2である。F1世代では全ての対立遺伝子がヘテロ接合型のため現れなかった表現型が、BC1F2ではどちらかの親型のホモ接合型で現れる。この原理を利用して、ゲノム上の多数の遺伝子変異から目的の変異を探し出すことができる。
6.ゲノム再解読
網羅的遺伝子発現解析で使われた次世代シーケンス技術をゲノムDNAに適用したもの。数億~数十億のゲノムDNA塩基数の数十倍に及ぶ大量の短い断片量を取得し、本来のゲノム配列と相互比較することで、各ゲノムが示す塩基配列上の違いを収集する。
7.ゲノム編集法
ゲノム上の特異的な塩基配列だけを標的とし、その近くに塩基の添加・欠落・置換などを行う実験手法。これまで複数の方法が開発されていて、近年ではCRISPRと呼ばれる分子ハサミを活用した編集法が、その精度と利便性から広く使われている。本研究でも、CRISPR法によりTAF12b遺伝子を破壊した。
8.転写補助因子
単独では転写活性を示さないが、他の転写因子の機能を補助する役割を持つ転写因子。
9.持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標。持続可能な世界を実現するための17のゴール、169のターゲットから構成され、発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に取り組んでいる(外務省ホームページから一部改変して転載)。
共同研究グループ
理化学研究所 環境資源科学研究センター
機能開発研究グループ
研究員(研究当時) キム・ジュンシク(June-Sik Kim)
(現 バイオ生産情報研究チーム 研究員)
上級研究員(研究当時) 高橋 史憲(たかはし ふみのり)
(現 客員研究員、東京理科大学 先進工学部 生命システム工学科 准教授)
研究員(研究当時) 浦野 薫(うらの かおる)
(現 客員研究員、農研機構 生物機能利用研究部門 契約研究員)
グループディレクター 篠崎 一雄(しのざき かずお)
細胞機能研究チーム
研究員 柴田 美智太郎(しばた みちたろう)
大阪大学大学院 理学研究科 生物科学専攻
助教 坂本 勇貴(さかもと ゆうき)
東京大学 大学院新領域創成科学研究科 先端生命科学専攻
教授 松永 幸大(まつなが さちひろ)
東京農業大学 農生命科学研究所
教授 篠崎 和子(しのざき かずこ)
研究支援
本研究は日本学術振興会(JSPS)科学研究費助成事業基盤研究(C)「ストレス応答欠如による根の伸長阻害を回復した突然変異植物の遺伝子機能解析(研究代表者:キム・ジュンシク)」、理化学研究所基礎科学特別研究員事業研究課題「植物の小胞体ストレス応答性転写因子による植物発生における機能解析(研究者:キム・ジュンシク)」による支援を受けて行われました。
原論文情報
June-Sik Kim, Yuki Sakamoto, Fuminori Takahashi, Michitaro Shibata, Kaoru Urano, Sachihiro Matsunaga, Kazuko Yamaguchi-Shinozaki and Kazuo Shinozaki, “Arabidopsis TBP-ASSOCIATED FACTOR 12 ortholog NOBIRO6 controls root elongation with unfolded protein response cofactor activity”, Proceedings of the National Academy of Sciences of the United States of America, 10.1073/pnas.2120219119
発表者
理化学研究所
環境資源科学研究センター 機能開発研究グループ
研究員(研究当時) キム・ジュンシク(June-Sik Kim)
(現 バイオ生産情報研究チーム 研究員)
グループディレクター 篠崎 一雄(しのざき かずお)
大阪大学大学院 理学研究科 生物科学専攻
助教 坂本 勇貴(さかもと ゆうき)
東京大学 大学院新領域創成科学研究科 先端生命科学専攻
教授 松永 幸大(まつなが さちひろ)
東京農業大学 農生命科学研究所
教授 篠崎 和子(しのざき かずこ)
報道担当
理化学研究所 広報室 報道担当
大阪大学 理学研究科 庶務係
東京大学 大学院新領域創成科学研究科 広報室
学校法人東京農業大学 経営企画部

