2023-08-09 東京大学
ドル 有生(奈良先端科学技術大学院大学 研究員/研究当時:生物科学専攻 博士課程)
古賀 皓之(生物科学専攻 助教)
塚谷 裕一(生物科学専攻 教授)
発表のポイント
- 葉にある気孔の作り方が植物種によって多様である仕組みに関する仮説を、気孔を作る遺伝子のはたらくタイミングに注目して証明した。
- 気孔をつくる遺伝子がはたらくタイミングを遺伝子操作によりずらすことで、気孔の作り方を思い通りに改変することに成功した。
- 植物の暮らしを支える気孔の進化過程の解明や、農業応用が期待される気孔の構造改変に向けて重要な知見が得られた。

遺伝子操作したシロイヌナズナの気孔(左)とベゴニアの気孔クラスター(右)
発表概要
東京大学大学院理学系研究科のドル有生大学院生(研究当時、現在は奈良先端科学技術大学院大学 日本学術振興会特別研究員)、古賀皓之助教、塚谷裕一教授による研究グループは、植物の気孔の作り方に多様性が生まれる仕組みを明らかにしました。
気孔は植物の葉の表面にある微小な穴で、光合成や呼吸に必要な気体を取り込む重要な役割を担います。気孔は一般的に気孔のもととなる細胞(気孔幹細胞)(注1)が分裂を重ねてから作られます。この作り方には種によって違いがあることが知られていました。例えば、気孔幹細胞が気孔になるまでに分裂する回数は種によってさまざまです。しかし、このような多様性が生まれる進化的な仕組みは分かっていませんでした。
研究グループは以前、オオバコ科のアワゴケ属(注2)というグループの植物で、気孔のもととなる細胞の分裂回数に多様性があることを見出し、その多様性が気孔を作るのに重要な遺伝子のはたらくタイミングの違いと相関することを明らかにしました。しかし、本当にそのタイミングの違いが、気孔の作り方に変化をもたらすのか否かは証明していませんでした。
そこで本研究では、この遺伝子のはたらくタイミングの違いを人為的に再現することで、実際に気孔の作り方に変化が生じるかを、実験モデル植物のシロイヌナズナ(注3)で検証しました。その結果、遺伝子のはたらく時期をずらす操作を行なった植物は、期待通りの気孔形成パターンを示すことが分かりました(図1)。
この結果から、気孔の作り方が種によって異なる要因として、気孔をつくる遺伝子のはたらくタイミングの変化があることを実証できました。この成果は、多様な気孔パターンの進化の仕組みを明らかにするとともに、気孔形態の人為的な改変の可能性を示すものとして重要です。

図1:研究の概要図
気孔をつくる際は、気孔(赤色)のもととなる気孔幹細胞(緑色)の分裂を維持する遺伝子SPCHと、その分裂を終結する遺伝子MUTEが順番にはたらくことが知られていた。本研究では、このSPCHとMUTEのはたらく時間間隔の違いが気孔の作り方の多様性を生み出しているのではないかと考え、シロイヌナズナにおける遺伝子操作によってMUTEのはたらくタイミングを変更し、実際に気孔の作り方が変化するかを調べた。その結果、予想通りMUTEを早くした時は気孔幹細胞の分裂が減り、MUTEを遅くした時は分裂が多くなり、それぞれシロイヌナズナとは異なる他の野生植物のようなパターンを示した。これにより、遺伝子のはたらく間隔に基づく、気孔作りの多様化の仕組みが実証された。
発表内容
〈研究の背景〉
植物は、葉の表面に気孔と呼ばれる小さな穴をもち、これを通して光合成や呼吸に必要な二酸化炭素や酸素を取り込んでいます。顕微鏡で色々な植物の気孔を眺めてみると、ある種では気孔がいつも3つの細胞に囲まれていたり、また他の種では2つの細胞に囲まれていたりと、気孔の並び方はさまざまなことが分かります(図2A)。実は、この見た目の違いは、気孔ができる過程の違いを反映したものです(図2B)。気孔は、葉の表面にある気孔のもととなる細胞(気孔幹細胞)が何回かの分裂を経て、最後に1組の孔辺細胞を生み出すことで作られます。この気孔幹細胞の分裂回数や分裂の方向などが、最終的な気孔の並び方を決める一因となっているのです。例えば気孔幹細胞の分裂がおこらないと、気孔は不規則型と呼ばれる特に法則性のない配置となりますが、平行に分裂すると2つの細胞に囲まれた平行型、らせん状に分裂すると3つの細胞に囲まれた不等型、そこからさらに分裂を繰り返すと、らせん型と呼ばれる並び方になります。ではこの気孔幹細胞の分裂回数は、どのように決まっているのでしょうか。
研究グループは以前、オオバコ科のアワゴケ属というグループの植物で、気孔のでき方に多様性があることを見出しました(2021年3月30日プレスリリース「植物の「暮らし」によって気孔の作り方が違う仕組みを解明」)。アワゴケ属では、各種の生態の違いと対応して、ある種では気孔幹細胞が分裂せずに直接気孔になるのに対し、他の種では気孔幹細胞が複数回分裂してから気孔になる、という違いがありました。また、両者の間では、気孔幹細胞の分裂を促進するはたらきを持つ遺伝子SPEECHLESS (SPCH)と、分裂を停止するはたらきを持つ遺伝子MUTEのはたらくタイミングが異なることも見出し、これが気孔の作り方の違いを生み出している原因ではないかという仮説をたてました。つまり、分裂を促進するSPCHと停止するMUTEがほぼ同時にはたらくと、気孔幹細胞は分裂せずに直接気孔になるが、MUTEのはたらくのが相対的に遅れた種では、気孔幹細胞が分裂してから気孔をつくるようになる、という仮説です。しかし、アワゴケ属の植物は遺伝子操作法が確立しておらず、この仮説を実証することはできませんでした。
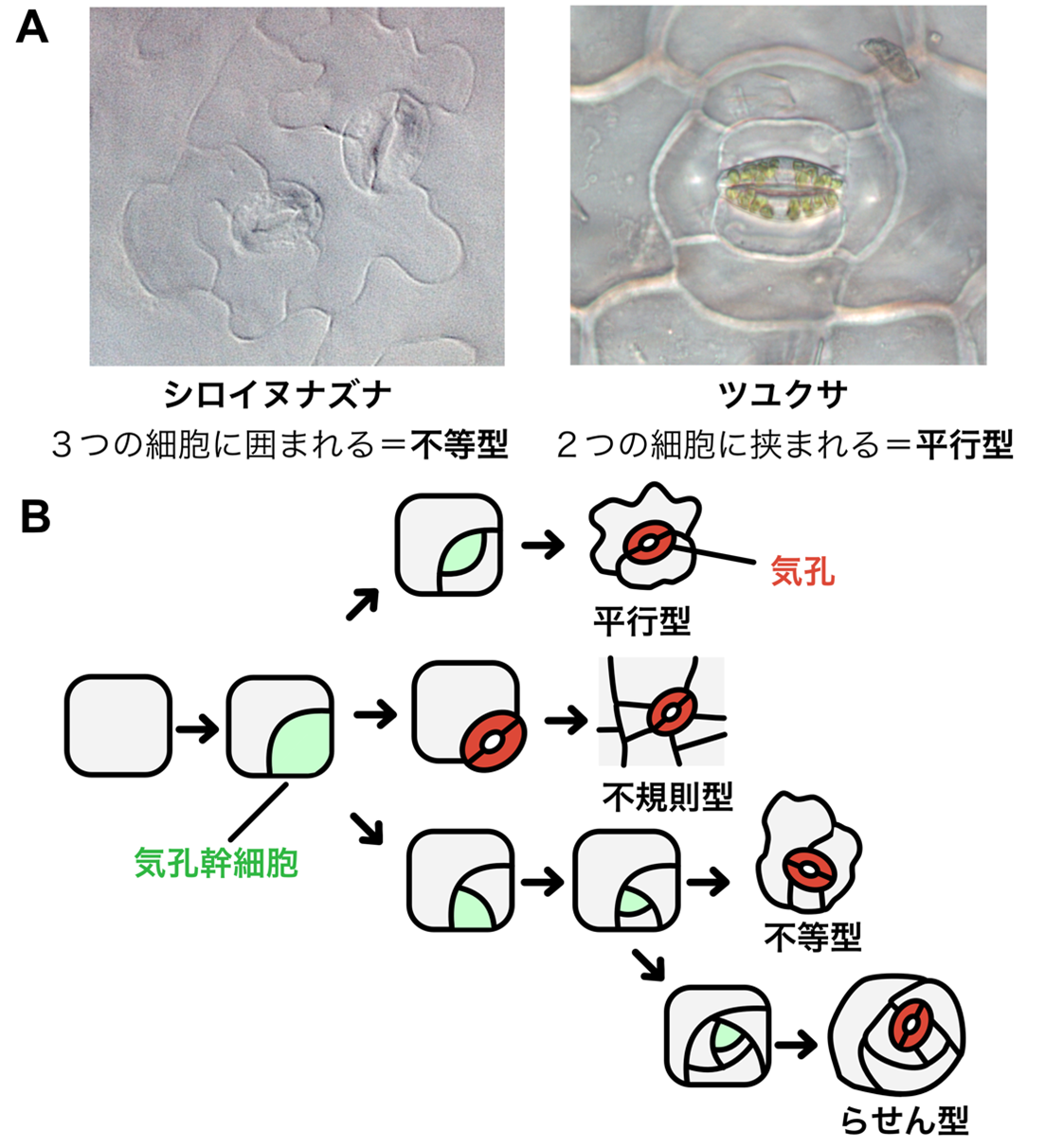
図2:気孔の並び方と気孔の作り方の関係
(A) シロイヌナズナとツユクサの気孔。両種では気孔を囲む細胞の数が異なっている。このように、植物の種によって気孔の並び方はさまざまである。(B) 野生植物にみられる気孔の並び方のパターン(右)と、それを生み出す気孔の作り方のパターン(左)。気孔幹細胞が気孔になるまでに、どのような方向に、何回分裂するかによって、最終的な気孔の並び方(「型」)が異なる。
〈研究の内容〉
本研究では遺伝子操作の簡単な実験モデル植物のシロイヌナズナを用いて、この仮説の検証を行ないました。シロイヌナズナの気孔は、気孔幹細胞が複数回分裂した後に作られます。つまり、仮説に基づけばSPCHがはたらいたあとMUTEがはたらくまでにやや間隔があると想定されます。ここでSPCHとMUTEのはたらく間隔を通常より短くしたり、長くしたりした時に、気孔幹細胞の分裂回数がそれぞれ減少または増加するかどうかを調べることで、上記の関係が成り立つかを検証できます(図1)。本研究では、特にMUTEのはたらくタイミングを操作することで検証を行いました。具体的にはまずMUTE遺伝子が壊れた変異体(注4)シロイヌナズナを準備し、これに通常と異なるプロモーター(注5)の制御下においたMUTE遺伝子を改めて導入しました。これによって、本来とは異なるタイミングでMUTEがはたらく遺伝子組み換え植物が得られます。実際には通常のMUTEよりも遅く発現する遺伝子のプロモーターを用いることで、MUTEがはたらくのが遅れた植物を、一方SPCHとほぼ同じタイミングではたらく遺伝子のプロモーターを用いることで、MUTEが早期にはたらく植物を得ることに成功しました(図1)。
次に、これら遺伝子組み換え植物における気孔のでき方を詳しく調べました(図3)。MUTEが遅れる植物では、気孔幹細胞の分裂回数の増加がみられたほか、らせん型の野生植物が示すのに似た、気孔と小さな細胞が密集した構造(注6)も観察されました。このことから、SPCHとMUTEのはたらく時間間隔の拡大が、気孔幹細胞の分裂回数の増加、ひいては気孔の並び方の変化に繋がりうることが示されました。一方、MUTEがはたらくのが早期化する植物では、気孔が隣り合い、一箇所に固まってできてしまう異常な構造が見られました。しかし、そうした異常を示さない気孔について調べると、通常よりも分裂回数が少ないまま形成された気孔が多いことが分かりました。これはMUTEの発現を早めることで、気孔幹細胞の分裂回数が減ったことを示しています。すなわち、MUTEを早くする実験、遅くする実験どちらについても、当初の仮説が支持されました。
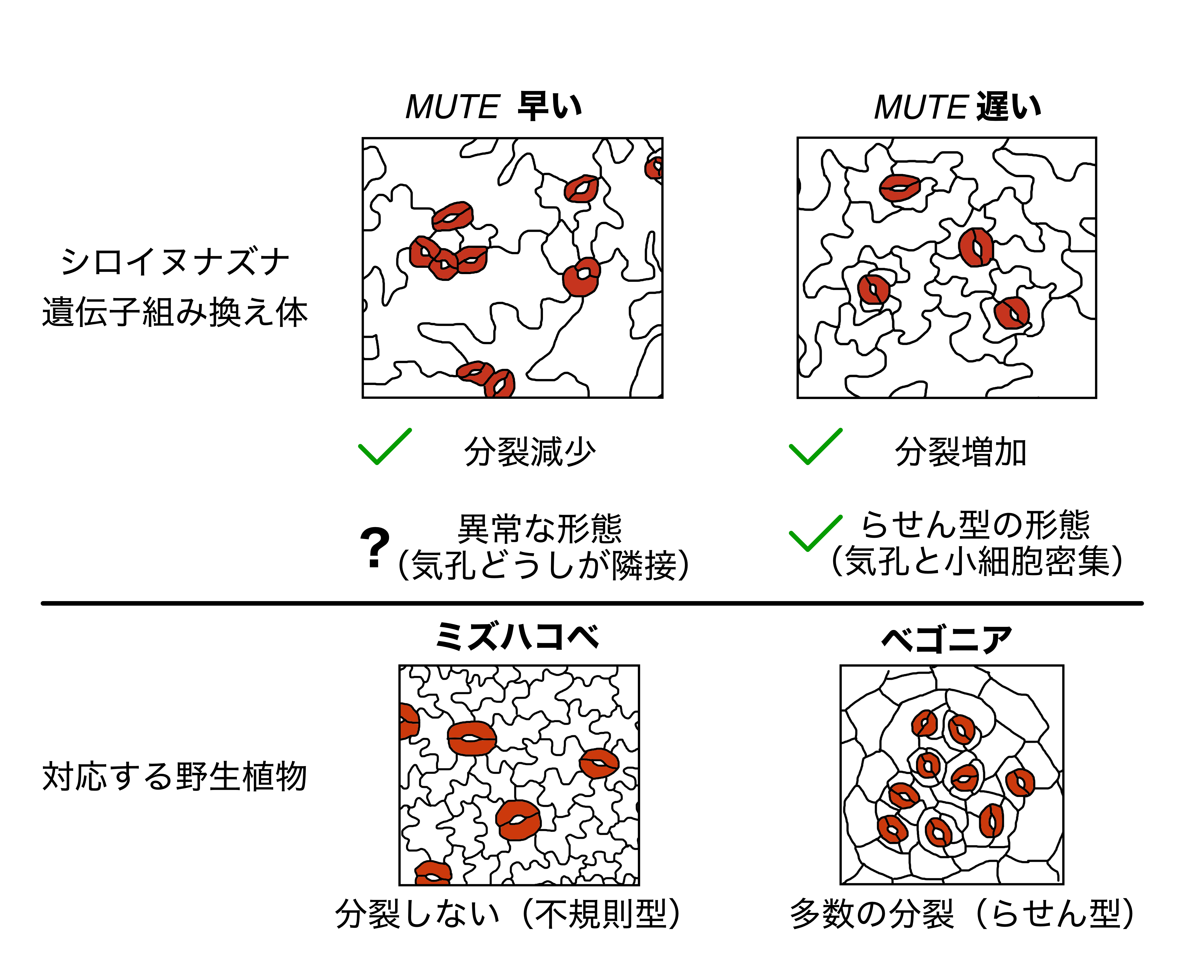
図3:本研究で作出された気孔の作り方が改変されたシロイヌナズナ
MUTEのはたらくのを早く、または遅くした遺伝子組み換え植物の気孔の形態を上段に示し、改変の目標となるシロイヌナズナとは異なる気孔の作り方をする野生植物の形態を下段に示した。それぞれの遺伝子組み換え植物では予想通り気孔幹細胞の分裂が減少、または増加した。MUTEを遅くした植物では分裂の多いらせん型の植物にみられる、気孔と小さな細胞が密集した構造が見られた。MUTEを早くした植物では気孔どうしが隣接する形態が見られたが、これは野生植物には見られない異常な形態である。
〈今後の展望〉
植物の気孔発生パターンの多様性は古くから認められていましたが、そうした多様性が生まれる仕組みは実証されていませんでした。本研究では、気孔作りを制御する遺伝子のはたらくタイミングの変化が、実際に気孔形成パターンの変化につながることを示し、これが気孔形成の多様化の仕組みになりうることを実証しました。これにより、植物の進化過程の理解や、光合成などをコントロールする上で農業的に重要な気孔形質の改良法の開発にむけて重要な知見が得られました。ただし今回の成果は、実際の進化を完全に模したものではなく、あくまで便宜的な操作による単純化された実験の結果であることには注意が必要です。例えばアワゴケ属では、今回MUTEのはたらきを早期化した植物で見られたような気孔形態の異常は伴わずに、気孔幹細胞の分裂の消失が起こっています。実際の植物種間における進化の直接の原因を明らかにするのが今後の課題です。
論文情報
- 雑誌名
Journal of Experimental Botany論文タイトル
Experimental validation of the mechanism of stomatal development diversification著者
Yuki Doll, Hiroyuki Koga*, Hirokazu TsukayaDOI番号
10.1093/jxb/erad279
研究助成
本研究は、「笹川科学研究助成(課題番号: 2019-4112)」、科研費「特別研究員奨励費(課題番号:20J20446)」、「若手研究(課題番号:20K15816)」、新学術領域研究 「植物の周期と変調(課題番号:19H05672)」の支援により実施されました。
用語解説
注1 気孔幹細胞
メリステモイドとも呼ばれ、多角形の形態を示す。複数回の分裂を経たのちに丸みを帯びた細胞(孔辺母細胞と呼ばれる)へと変化し、その後1回分裂することで気孔を構成する孔辺細胞となる。
注2 アワゴケ属
コケとつくがコケの仲間ではなく、花を持つ被子植物である。オオバコ科に属し、世界中に50種以上が生息する植物の属。水中と陸上の両方で生育できる両生種から、陸上でのみ生育する陸生種、水中でのみ生育する水生種まで多様な生態の種を含む。本研究グループはアワゴケ属を対象に進化生物学的研究を行なってきた(2021年3月30日プレスリリース「植物の「暮らし」によって気孔の作り方が違う仕組みを解明」、2021年7月29日プレスリリース「水陸両生の水草ミズハコベが姿を変える仕組みを解明」などを参照)
注3 シロイヌナズナ
世界中でもっともよく分子生物学・遺伝学的な実験に用いられている植物(モデル植物)で、アブラナ科の一年草。気孔の発生の研究はシロイヌナズナを用いて大きく進んできた。遺伝子操作の技術がよく発達しているため、簡単に外来遺伝子の導入ができる。
注4 変異体
特定の遺伝子の配列に変化(変異)が生じ、その遺伝子の機能に変化が生じた生物。本研究の場合、MUTE遺伝子に変異が生じたことにより、MUTE遺伝子の機能が完全に失われた変異体(機能欠損変異体)を用いた。
注5 プロモーター
遺伝子の近傍にあって、その遺伝子のはたらきを調節する役割をもつDNA領域。ここではとくに、その遺伝子のはたらきの時空間的なパターンを再現するのに必要な領域全体を指す。本研究で用いた遺伝子については、過去の研究でプロモーターが特定されている。
注6 気孔と小さな細胞が密集した構造
気孔クラスターとも呼ばれる。気孔幹細胞が局所的に分裂を多数回行なうことによって形成される構造であり、ベゴニアやユキノシタなど、らせん型の気孔を持つ植物に特徴的にみられる。図3右下のベゴニアは典型的な気孔クラスターを示しており、図3右上の遺伝子操作をしたシロイヌナズナはそれに似た構造を示している。
")
")