2024-03-22 理化学研究所,名古屋大学,横浜市立大学,神戸大学,京都先端科学大学,京都大学
理化学研究所(理研)環境資源科学研究センター 代謝システム研究チームの鵜崎 真妃 基礎科学特別研究員、平井 優美 チームリーダー(名古屋大学大学院 生命農学研究科 客員教授)、横浜市立大学 理学部 理学科の山本 浩太郎 助教、京都先端科学大学 バイオ環境学部 バイオサイエンス学科の三村 徹郎 教授、神戸大学大学院 理学研究科 生物学専攻の石崎 公庸 教授、深城 英弘 教授、京都大学大学院 理学研究科 生物科学専攻の大西 美輪 博士研究員らの国際共同研究グループは、薬用植物ニチニチソウ[1]の種子胚[2]におけるアルカロイド[3]生合成開始過程を明らかにし、アルカロイド代謝において細胞分化が重要な役割を担う可能性を示しました。
本研究成果は、抗がん剤などの薬として重要なニチニチソウアルカロイドの生合成およびその制御機構の理解につながり、植物や植物細胞を用いた化合物生合成技術の開発に貢献すると期待されます。
植物は多様な特化代謝[4]産物を合成・蓄積し、その中には薬や嗜好(しこう)品として人間生活において重要な役割を担っているものが多くあります。特化代謝の多くは細胞種特異的に行われることが知られていますが、その理由は未解明です。今回、国際共同研究グループは、抗がん剤などの薬として有用なアルカロイドを細胞種特異的に合成するニチニチソウの種子胚において、種子発芽に伴う細胞の状態変化、アルカロイド生合成の開始過程やアルカロイドの細胞局在を明らかにしました。本研究は、科学雑誌『New Phytologist』オンライン版(3月22日付:日本時間3月22日)に掲載されました。

ニチニチソウ種子の成熟・発芽過程におけるアルカロイド生合成の開始時期を解明
背景
自らは移動しない植物は、昆虫や草食動物、病原菌などの外敵への防御のため、また周囲の環境に適応するため、特化代謝産物と呼ばれる化合物を合成・蓄積しています。特化代謝産物は、特定の植物種のみが特定の環境の下で合成することが多く、人間生活では薬や嗜好品として重要な役割を持つものが数多く存在します。これらの特化代謝産物は非常に複雑な化学構造を持つため、人工合成が難しく、植物からの抽出に頼っています。一方、有用な特化代謝産物は希少植物や生育の極めて遅い植物種がごく少量しか合成しない場合が多いのが現状です。そのため、植物がこれらの化合物を合成する仕組みや制御機構を知ることは、植物による合成量を増加させたり、酵母や大腸菌といった工業的な培養の容易な生物に合成させる手法を開発したりする上で重要です。
薬用植物であるニチニチソウは、抗がん剤として使われるビンブラスチンやビンクリスチンをはじめとする多種多様なアルカロイドを合成します。ニチニチソウのアルカロイドは、葉では複数の異なる種類の細胞中で、多段階の酵素反応を経て合成されます。まず、IPAP(Internal Phloem Associated Parenchyma)細胞と呼ばれる細胞で合成された前駆体が表皮細胞へ運搬されます。表皮細胞でさらに多段階の反応を経て中間代謝物に変換された後、異形細胞・乳管細胞と呼ばれる細胞へと運搬されて最終産物であるアルカロイドとなって蓄積されます。なぜニチニチソウアルカロイド生合成がこのような複雑な工程からなるのかは不明です。生合成経路のうちの特定の酵素反応が進行するためには、反応の場である細胞が特定の性質を持っている必要があるのかもしれません。
国際共同研究グループは、特定の性質を持たない幹細胞が、異なる性質を持つ各種細胞に変化する「分化」に注目しました。そして、植物の「形態的な」分化がおおむね完了した上で休眠状態になっている種子を研究対象としました。乾燥した種子では代謝が停止していますが、水を与えると代謝を再開して発芽します。国際共同研究グループは、ニチニチソウの種子発芽過程において、アルカロイド蓄積量と生合成関連遺伝子の発現量、さらにアルカロイドの主な蓄積場所である乳管細胞の形態的変化を調べることで、細胞が「代謝的に」分化する過程の解明に挑みました。
研究手法と成果
国際共同研究グループは、受粉後の成熟過程にある種子、完熟後に吸水させた種子、そして発芽過程の種子から胚を取り出して光学顕微鏡および蛍光顕微鏡で観察し、乳管細胞が成熟途上の種子の胚においてすでに形態的に分化していることを発見しました。発芽過程の胚の細胞を電子顕微鏡で詳細に観察したところ、乳管細胞周辺の細胞では、一般的に発芽前・直後の胚で観察される構造が見られた一方で、乳管細胞では分解型液胞と呼ばれる細胞小器官が発達しているなど、細胞内構造が周囲の細胞とは大きく異なっていました(図1)。

図1 種子胚切片の顕微鏡写真
上:成熟前の種子から取り出した胚の切片の光学顕微鏡写真。この時期にすでに細長い乳管細胞(赤破線で囲んだ部分)が形態的に分化していた。
下:発芽24時間後の種子から取り出した胚の切片の光学顕微鏡写真および電子顕微鏡写真。乳管細胞と周囲の皮層細胞では細胞内構造が異なっている。スケールバーは0.5mmを表す。
そこで、次に完熟後の種子に着目し、吸水前、吸水後発芽前、発芽後の胚を取り出してアルカロイド蓄積量と遺伝子発現量の変化を調べた結果、アルカロイド生合成は発芽後12時間目以降に活発になることが分かりました。アルカロイド生合成酵素遺伝子の発現パターンを見ると、発芽後12~24時間目から発現量が大きく増加する遺伝子と、発芽後36時間目から発現量が増加する遺伝子とに分かれました。葉においては、生合成酵素遺伝子がどの細胞で発現するのかが明らかになっています。種子胚において発芽後12~24時間目、および36時間目に発現量が増大する遺伝子は、葉において、前者は表皮細胞、後者は異形細胞、乳管細胞およびIPAP細胞で発現していました(図2)。
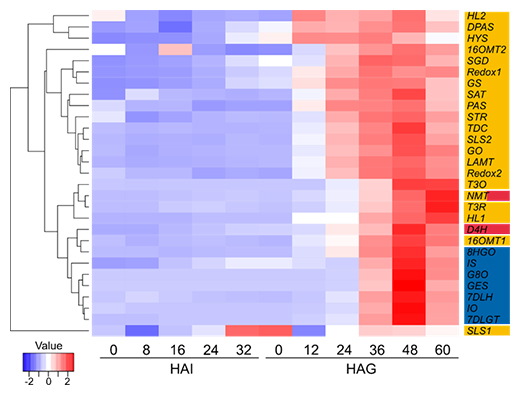
図2 アルカロイド生合成酵素遺伝子の種子吸水・発芽に伴う発現変動
種子吸水開始から0~32時間後(HAI)および発芽0~60時間後(HAG)のアルカロイド生合成酵素遺伝子の発現変動を表すヒートマップ。色はデータを標準化(各データ値から平均値を引いた値を標準偏差で割ること)した各遺伝子の各時点での発現量を表している。葉において、表皮細胞で発現することが知られている遺伝子は黄色、異形細胞・乳管細胞で発現することが知られている遺伝子は赤色、IPAP細胞で発現することが知られている遺伝子は青色の背景で示した。
葉では異形細胞と乳管細胞にのみアルカロイドが蓄積するため、成長途上の種子胚でも同様かを確認しました。アルカロイドと反応し褐色沈殿を作るドラーゲンドルフ試薬で発芽後48時間目の胚の切片を染色したところ、乳管細胞が褐色に染色され、種子胚においても乳管細胞特異的にアルカロイドが高蓄積していることが示されました(図3)。
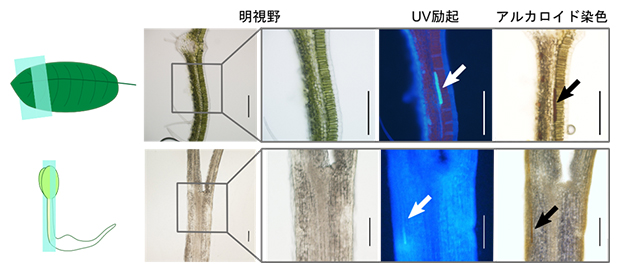
図3 葉および種子胚の切片のアルカロイド染色
アルカロイドと反応して褐色沈殿を作るドラーゲンドルフ試薬を用いて染色した、葉(上)および種子胚(下)の切片の光学顕微鏡写真。UV励起すると自家蛍光を発する乳管細胞が茶色に染色されている。スケールバーは0.2mmを表す。
これまで、特化代謝産物の機能は成長した植物体が外敵から身を守ることであると考えられてきたため、多くの研究は成熟した葉を用いて行われてきました。種子に着目した本研究の成果は、植物の一生の中で最も脆弱(ぜいじゃく)な時期といえる発芽から芽生えの時期においても、アルカロイドはニチニチソウにとって重要な役割を担っていることを示唆しています。
また、ニチニチソウのアルカロイドは虫や草食動物からの被食に応答して合成されることが知られていましたが、今回の研究から、植物の発生・成長にも連動して合成されることが明らかになりました。ニチニチソウアルカロイド生合成には30もの合成酵素遺伝子が働いています。これらの遺伝子が発芽後一斉に活性化されるのではなく、発生段階や発現場所に関連した異なる制御を受けていることには生理学的な理由があると考えられます。
今後の期待
今回の研究により、発生・成長に連動したニチニチソウアルカロイド合成の制御機構の存在が示唆されました。今後、この未知の制御機構を研究することで、ニチニチソウが非常に複雑な化学構造を持つアルカロイドを合成・蓄積できる仕組みの解明につながります。
一般に、植物の特化代謝産物は複雑な化学構造を持ち人工的な合成が難しい一方で、人間生活において重要な役割を担うことから、その安定的な生産が求められています。植物特化代謝を酵母など培養の容易な生物で再現し、効率的に植物特化代謝産物を合成しようとする研究が世界中で行われていますが、いまだ高効率での化合物生合成には成功していません。知られざる巧みな特化代謝とその制御機構を植物から学ぶことは、人間にとって有用な植物特化代謝産物の安定供給を目指す研究の発展に大いに貢献するものです。
今回の研究は、国際連合が定めた17項目の「持続可能な開発目標(SDGs)[5]」のうち、「3.すべての人に健康と福祉を」への貢献が期待できます。
補足説明
1.ニチニチソウ
マダガスカル原産のキョウチクトウ科植物で、300種類以上の多様なアルカロイドを合成・蓄積する。ニチニチソウの合成するアルカロイドには、抗がん剤として使用されるビンブラスチンやビンクリスチンなど、人間生活において重要な役割を持つ化合物が多く含まれるため、その生合成経路がよく研究されている。
2.種子胚
種子中に含まれる、将来植物体になる部分のこと。
3.アルカロイド
ラテン語で「アルカリのようなもの」という意味で、多くが塩基性を示す。厳密な定義は難しいが、一般的には窒素原子を含む低分子化合物のうち、アミノ酸や核酸など他のカテゴリーに分類されるものを除いたものを指す。モルヒネやニコチンなど、生理活性を持つものが多く知られている。
4.特化代謝
植物種に固有の代謝。二次代謝とも呼ばれる。
5.持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標。持続可能な世界を実現するための17のゴール、169のターゲットから構成され、発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に取り組んでいる(外務省ホームページから一部改変して転載)。
国際共同研究グループ
理化学研究所 環境資源科学研究センター
代謝システム研究チーム
基礎科学特別研究員 鵜崎 真妃(ウザキ・マイ)
チームリーダー 平井 優美(ヒライ・マサミ)
(名古屋大学 大学院生命農学研究科 客員教授)
質量分析・顕微鏡解析ユニット
専門技術員 森 哲哉(モリ・テツヤ)
技師 佐藤 繭子(サトウ・マユコ)
テクニカルスタッフⅡ 若崎 眞由美(ワカザキ・マユミ)
上級技師 豊岡 公徳(トヨオカ・キミノリ)
横浜市立大学 理学部 理学科
助教 山本 浩太郎(ヤマモト・コウタロウ)
京都先端科学大学 バイオ環境学部 バイオサイエンス学科
教授 三村 徹郎(ミムラ・テツロウ)
(神戸大学 大学院理学研究科 名誉教授)
神戸大学 大学院理学研究科 生物学専攻
准教授 村上 明男(ムラカミ・アキオ)
助教 七條 千津子(シチジョウ・チヅコ)
教授 石崎 公庸(イシザキ・キミツネ)
教授 深城 英弘(フカキ・ヒデヒロ)
京都大学 大学院理学研究科 生物科学専攻
博士研究員 大西 美輪(オオニシ・ミワ)
マックス・プランク化学生態学研究所(ドイツ)
ディレクター サラ・オコナー(Sarh E. O’Connor)
テクニカルスタッフ ダリア・アイレド・セレナ・グエロ(Delia Ayled Serna Guerrero)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業新学術領域研究(研究領域提案型)「環境感覚を支える植物液胞動態とその適応機構(研究代表者:三村徹郎、22120006)」「植物の力学的最適化戦略に基づくサステナブル構造システムの基盤創成(領域代表者:出村拓)」の計画研究「顕微技術を駆使した計測と制御による細胞構造のしなやかさの高精度解析(研究代表者:細川陽一郎、18H05493)」、同基盤研究(B)「細胞横断型植物二次代謝機構における細胞分化過程の解明(研究代表者:三村徹郎、16H04807)」、同若手研究「1細胞オミクスによる多様な植物異形細胞の機能解明(研究代表者:山本浩太郎、22K15136)」、科学技術振興機構(JST)革新的GX技術創出事業(GteX)「先端的植物バイオものづくり基盤の構築(研究代表者:大熊盛也、JPMJGX23B0)」、同次世代研究者挑戦的研究プログラム「東海国立大学機構融合フロンティア次世代研究事業(JPMJSP2125)」、理化学研究所若手研究者育成制度「大学院リサーチ・アソシエイト制度」による助成を受けて行われました。
原論文情報
Mai Uzaki, Tetsuya Mori, Mayuko Sato, Mayumi Wakazaki, Noriko Takeda-Kamiya, Kotaro Yamamoto, Akio Murakami, Delia Ayled Serna Guerrero, Chizuko Shichijo, Miwa Ohnishi, Kimitsune Ishizaki, Hidehiro Fukaki, Sarah E. O’Connor, Kiminori Toyooka, Tetsuro Mimura, Masami Yokota Hirai , “Integration of cell differentiation and initiation of monoterpenoid indole alkaloid metabolism in seed germination of Catharanthus roseus”, New Phytologist, 10.1111/nph.19662
発表者
理化学研究所
環境資源科学研究センター 代謝システム研究チーム
基礎科学特別研究員 鵜崎 真妃(ウザキ・マイ)
チームリーダー 平井 優美(ヒライ・マサミ)
(名古屋大学 大学院生命農学研究科 客員教授)
横浜市立大学 理学部 理学科
助教 山本 浩太郎(ヤマモト・コウタロウ)
京都先端科学大学 バイオ環境学部 バイオサイエンス学科
教授 三村 徹郎(ミムラ・テツロウ)
神戸大学 大学院理学研究科 生物学専攻
教授 石崎 公庸(イシザキ・キミツネ)
教授 深城 英弘(フカキ・ヒデヒロ)
京都大学 大学院理学研究科 生物科学専攻
博士研究員 大西 美輪(オオニシ・ミワ)
報道担当
理化学研究所 広報室 報道担当
東海国立大学機構 名古屋大学広報課
横浜市立大学 広報課長 上村一太郎
神戸大学 総務部広報課
京都先端科学大学 広報センター
京都大学 渉外部広報課国際広報室
")
