2020-02-27 東京大学
木村 遼希(生物科学専攻 博士課程1年生)
橋本(杉本)美海(名古屋大学大学院生命農学研究科 助教)
射場 厚(九州大学大学院理学研究院 生物科学部門 教授)
寺島 一郎(生物科学専攻 教授)
矢守 航(現:大学院農学生命科学研究科 附属生態調和農学機構 准教授
/研究当時:生物科学専攻 准教授)
発表のポイント
- 野外では、雲の動きや上部に存在する葉の動きによって葉の受ける光強度は大きく変動する。光強度が一定の環境で光合成能力を強化した例はいくつかあるが、「変動する光環境」に対する植物の光合成応答の強化に成功した研究例はまだ極めて少ない。
- 気孔は環境に応じて開閉することによって、光合成に必要な二酸化炭素の取り込みや蒸散による水分の放出を制御している。本研究では、光強度の上昇に応じて気孔をすばやく開かせることで、野外の光環境を模した変動光環境において光合成および植物成長を促進することに成功した(図1)。
- 野外の変動する光環境における光合成の調節メカニズムの全貌の解明は、地球レベルの大気CO2の削減や食料増産のために必須な光合成効率の改善や光合成生産向上のための技術基盤となるだろう。

図1 : 気孔を迅速に開口させることで、野外における光合成応答と植物成長の促進に成功
発表概要
野外環境において、雲の切れ間から降り注ぐ光や、風で揺らめく植物の葉の間から差し込む光によって、植物の受ける光量は頻繁に変動している。弱光下に置かれた葉に強い光があたると光合成速度は徐々に上昇し、やがて定常状態に達する。この現象を光合成誘導反応(注1)と呼ぶ。光合成誘導反応が起こっている間、植物は本来の光合成能力を最大限発揮することができていない。そこで、光合成誘導に要する時間を短縮することによって、野外の変動する光環境における植物生産性を向上させようという取り組みが、世界中でなされている。
東京大学大学院理学系研究科の木村遼希 大学院生、大学院農学生命科学研究科の矢守航 准教授(研究当時:大学院理学系研究科 准教授)らは、モデル植物であるシロイヌナズナの気孔(注2)が開いたままの変異体では、野生型の個体に比べて光合成誘導期間が90%ほども短縮されることを見出した(図2)。
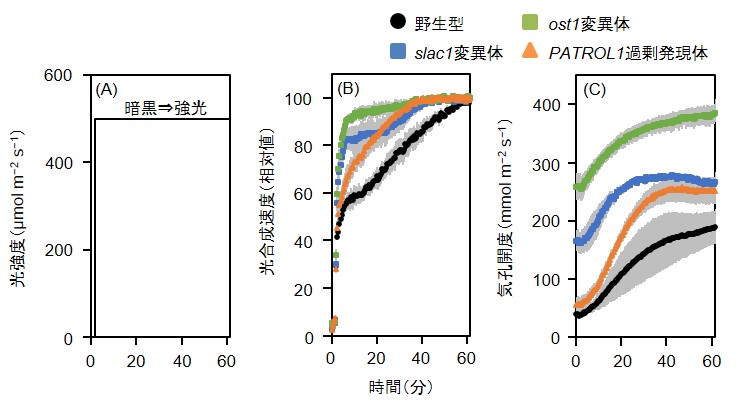
図2 : 高い気孔開度は光合成誘導に要する時間を短縮する
一晩暗黒下においたシロイヌナズナの葉に対して(A)のように突然強光を照射することで光合成誘導を起こし、ガス交換測定装置(GFS-3000)によって光合成速度(B)および気孔開度(C)を解析した。気孔開度は気孔の開き具合を表す数値で、大きければそれだけ気孔が開いていることを示す。気孔開度は絶対値で大きな差を示し、slac1変異体、ost1変異体は野生型より常に大きく気孔を開いていた。また、PATROL1過剰発現体は光照射後に野生型よりもすばやく気孔を開いた。光合成速度については定常状態の値を100とした相対値を算出したところ、定常状態に至るまでに要する時間に大きな違いが見られた。光合成速度が定常状態の70%の値に至るまでの所要時間を野生型とslac1変異体、ost1変異体、PATROL1過剰発現体との間で比較すると、slac1変異体とost1変異体では90%、PATROL1過剰発現体では65%短縮されていた。
この光合成誘導に要する時間の短縮によって、変動する光環境における光合成速度が最大で45%、植物体の成長量が50%増加した(図3、4、5)。
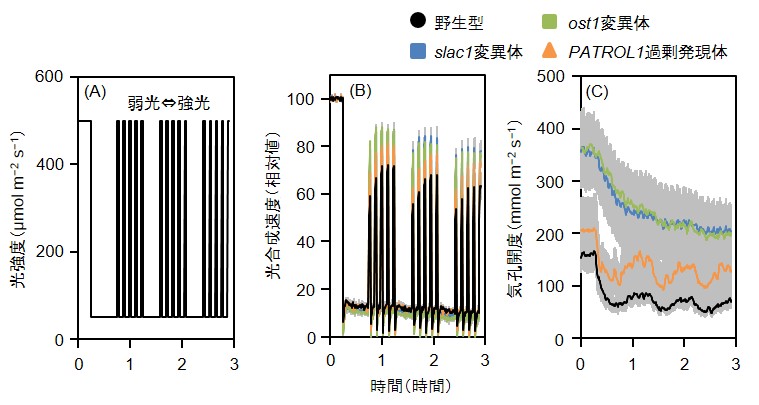
図3 : 高い気孔開度は変動光への光合成応答を向上させる
(A)のような人工的な変動光パターンを装置の光源で再現し、変動光に対する光合成応答(B)および気孔応答(C)をガス交換測定装置(GFS-3000)によって解析した。光合成速度については初めに葉に十分に強光を当てて得られた定常状態の値を100とし、相対値を算出した。気孔が大きく開いているslac1変異体、ost1変異体や、気孔がすばやく開口するPATROL1過剰発現体は、突然の強光に対しても野生型より定常状態に近い光合成速度を示した。
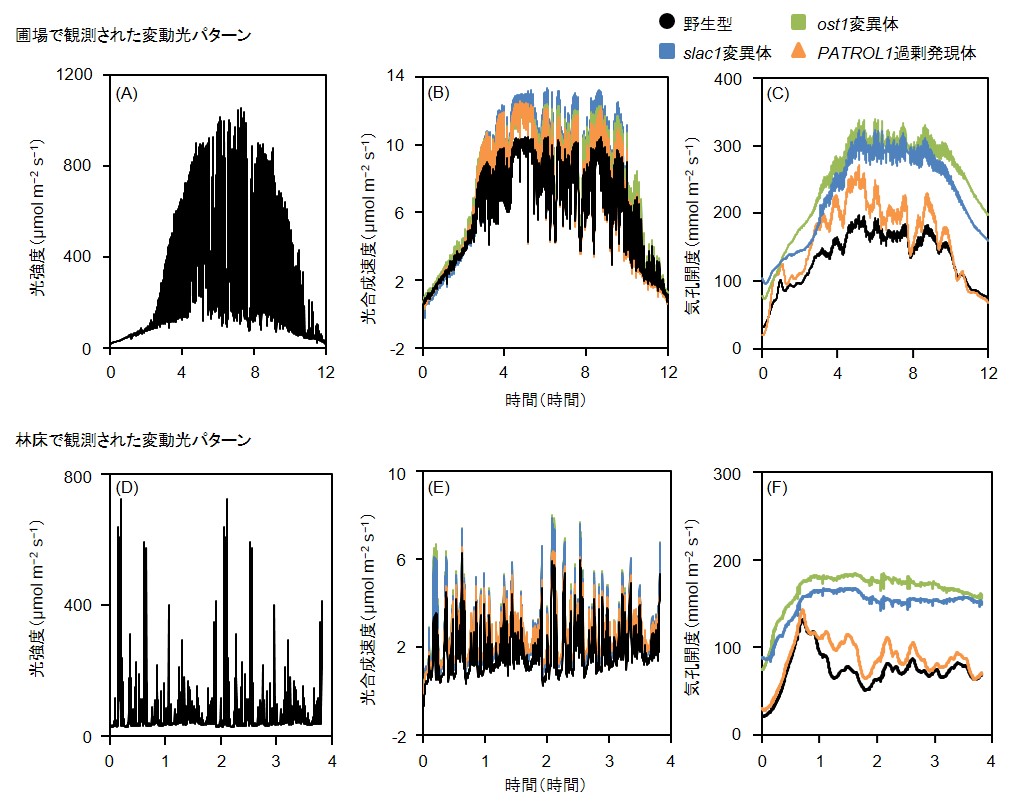
図4 : 気孔を大きく開くことで野外光条件においても光合成が促進される
(A)あるいは(D)のような、実際に野外環境で観測された変動光のパターンをガス交換光合成測定装置(Li-6400XT)の光源で再現し、一晩暗黒下においたシロイヌナズナの葉に照射して光合成速度(B、E)および気孔開度(C、F)を解析した。どちらの光条件でも、slac1変異体とost1変異体は常に野生型より大きい気孔開度を示し、PATROL1過剰発現体は野生型よりも変動光に対して機敏に応答していた。それに伴い、slac1変異体、ost1変異体、PATROL1過剰発現体では、光合成速度も野生型より大きかった。
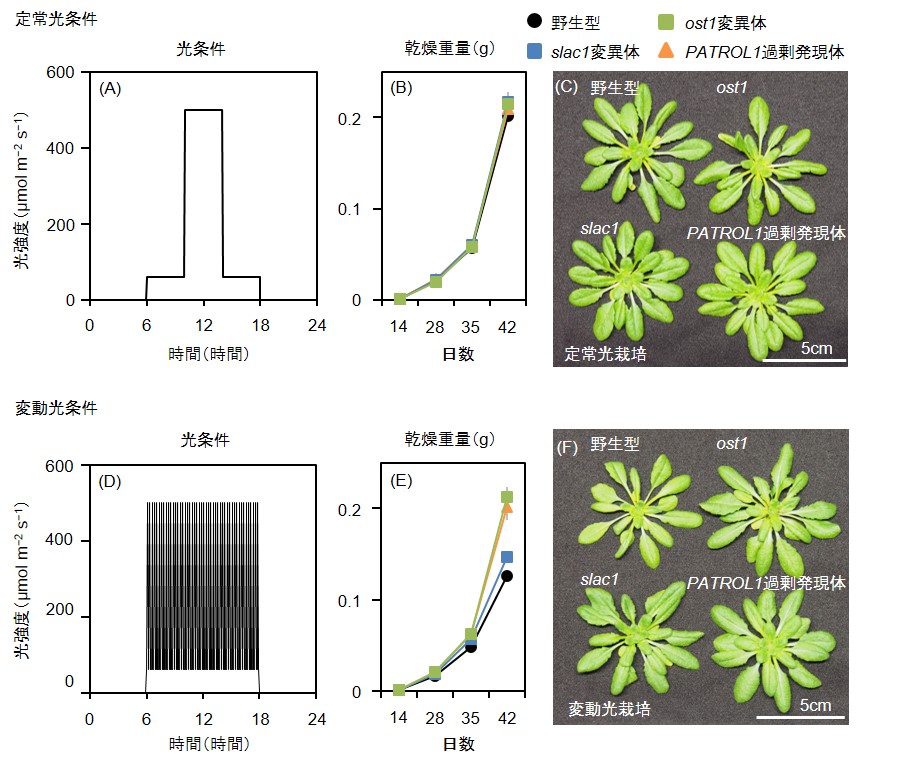
図5 : 高い気孔開度は変動光条件下での植物成長量を増加させる
播種後2週間まで定常光条件で育てた植物を、(A)のような定常光条件と(D)のような変動光条件とに移して栽培し、サンプリングして乾燥重量の推移を求めた(B、E)。定常光条件では差は見られなかったが、変動光条件ではost1変異体とPATROL1過剰発現体が野生型よりも有意に大きく成長した。(C、F)は播種後35日時点でサンプリングした植物の写真である。
しかし、これらの変異体では、気孔が開いていることでCO2を取り込むことはできるものの、同時に多くの水も失われるため、水利用効率(注3)が野生型に比べて著しく低かった(表1)。
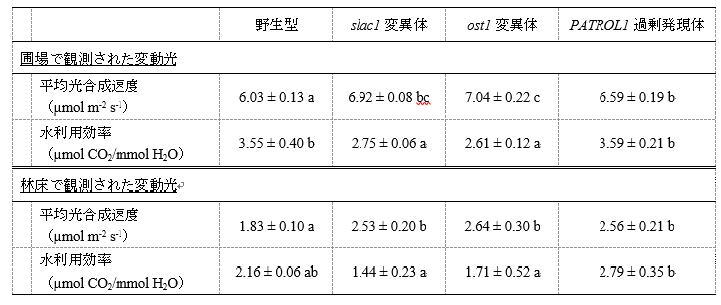
表1 : すばやい気孔開口を示すPATROL1過剰発現体は高い光合成速度と高い水利用効率を両立した
図4のデータをもとに、測定中の平均光合成速度と水利用効率を算出した。slac1変異体、ost1変異体、PATROL1過剰発現体のすべてが野生型よりも高い光合成速度を示していた。slac1変異体、ost1変異体の水利用効率は野生型と比較して低くなっていたが、PATROL1過剰発現体は野生型と同程度の水利用効率を示した。異なるアルファベットは有意差があることを示す。
そこで、気孔が光に応じてすばやく開閉する形質転換体を用いて同様の測定を行ったところ、光合成誘導期間は野生型に比べて65%短縮された(図2)。気孔開口の迅速化によって、変動する光環境における光合成速度が最大で40%、植物体の成長量は50%も増加した(図3、4、5)。さらに、特筆すべきは、この形質転換体は環境に応じて気孔を適切に閉鎖するので、野生型と同レベルの水利用効率を示したことである(表1)。一方、光強度が一定の定常光環境で植物を栽培すると、野生型とこれらの変異体や形質転換体との間に成長量の差はなかった(図5)。
本研究では、気孔開口状態の維持および気孔の光応答迅速化によって、光合成誘導に要する時間が短縮され、野外で見られるような変動光環境下において光合成速度および植物体の成長速度が向上することを実証した。さらに、気孔の光応答迅速化によって、野生型よりも高い光合成速度を示しながらも、野生型と同等の水利用効率を達成できることも明らかにした。最近、矢守 准教授らは、イネにおいても気孔開度の維持によって変動光環境下の光合成が増加することを見出した(Yamori et al. (2020) Plant, Cell & Environment)ため、小型モデル植物シロイヌナズナを用いた本研究の成果は、大型作物にも適用できる。これらの研究は、植物の気孔応答には最適化の余地があり、さまざまな変動環境に対する気孔応答の最適化によって野外環境における光合成速度の促進と作物生産性の強化が実現可能であることを強く示唆している。
発表内容
世界の人口増加はとどまるところを知らず、増大する食料需要に応え、将来にわたって食料を安定供給していくことは世界的に重要課題となっている。光合成は植物の成長量や収量を決定する最も重要な代謝であるため、高い光合成能力を有し、収量の高い作物の開発や利用は、食料生産を増大させる手段の一つとして注目されている。これまでにも光合成の機構解明やその改良を目指した研究は行われてきたが、その大部分は、光強度が一定の定常条件下における光合成に着目したものだった。農地や林床などでは、雲の切れ間から降り注ぐ光や、風で揺らめく植物の葉の間から差し込む光の影響で、植物の受ける光量は頻繁に変動している。植物は、これらの変動光環境下で効率よく光合成をおこなう必要がある。弱光下に置かれた葉に強光があたると、光合成速度は徐々に上昇し、やがて定常状態に達する。この現象を光合成誘導反応と呼ぶ。光合成誘導反応が起こっている間、植物は本来の光合成能力を発揮できていないことになる。また、暗黒下や弱光下では光合成関連酵素の不活性化や気孔の閉鎖が起こり、再度光合成誘導を必要とする状態になるため、野外環境では光合成誘導が頻繁に起こる。したがって、光合成誘導反応に要する時間を短縮することができれば、変動する光環境において、吸収した光エネルギーを有効利用でき、作物生産性の向上に大きく貢献すると考えられる。
光合成に必要なCO2は気孔から葉内に取り込まれる。しかし、気孔の開口は光の変動速度にくらべて遅いので、気孔葉内のCO2不足は光合成を大きく制限しうる。そこで、本研究では、気孔応答性の異なる植物がさまざまな光条件に対してどのような光合成応答を示すのかを解析した。使用した植物材料は、気孔を大きく開口する変異体であるslac1(slow anion channel-associated 1)変異体とost1(open stomata 1)変異体(注4)、CO2濃度の変化に対して気孔開口の素早いPATROL1(PROTON ATPase TRANSLOCATION CONTROL 1)過剰発現体(注5)である。まず、光合成活性測定装置(Dual-PAM)および高性能のガス交換光合成測定装置(GFS-3000, Li-6400XT)を用いて、強光照射による光合成誘導反応や、変動光環境下の光合成速度を比較した。また、栽培実験も行った。一定の光強度下では、これらの変異体と形質転換体の光合成速度や植物成長は、野生型と差がなかった(図5)。しかし、気孔が開いたままの変異体であるslac1変異体とost1変異体では、光合成誘導に要する時間が90%も短縮され(図2)、変動光に対する光合成応答も改善するほか(図3、4)、ost1変異体では、植物の成長量も野生型と比べて50%増加した(図5)。気孔では、光合成に必要なCO2の取り込みと同時に、蒸散による水分の放出が起こるので、気孔を大きく開くだけでは植物の水利用効率は低下し、乾燥ストレスを受けやすくなる。実際に、slac1変異体とost1変異体では、野生型に比べると水利用効率は顕著に低かった(表1)。一方で、PATROL1過剰発現体も、光照射に対して迅速な気孔開口を示し、光合成誘導に要する時間が65%短縮された(図2)。野外環境を模した変動光環境下では、光合成速度が最大で40%増加し(図3、4)、植物体の成長量も50%増加した(図5)。さらに、PATROL1過剰発現体では、環境に応じて気孔を適切に閉鎖することができるため、野生型と同じく高い水利用効率を示した(表1)。このように、本研究の結果は、植物の気孔応答には最適化の余地があり、さまざまな動環境に対する気孔応答の最適化によって、野外環境における光合成速度の促進と生産性強化が実現可能であることを示している。
植物の光合成応答は私たちの呼吸に必要な酸素を生産するだけでなく、農作物や化石燃料、その他のバイオマスを生産するエネルギー変換反応であるため、地球上のあらゆる生命にとって非常に重要な反応である。本研究を通して、野外の変動する光環境下の光合成や植物成長は、気孔の開きやすさに大きく制限されていることが明らかとなった。今回の成果は、野外環境での植物の収量増加を考える際に、気孔応答やそれに寄与するPATROL1が非常に有望なターゲットになることを示唆している。PATROL1遺伝子と非常に似た遺伝子がイネやソルガムなどの作物やポプラなどの樹木にも存在しているため、これらの植物においても過剰発現体によるバイオマス増産に期待できる。野外の変動する光環境における光合成の調節メカニズムの全貌解明は、光合成効率やバイオマス生産の向上のための技術開発のための基盤となり、地球レベルの大気CO2濃度上昇の抑制や食料増産に大きく貢献するはずである。
本研究成果は2月25日付でJournal of Experimental Botanyに掲載されました。
発表雑誌
- 雑誌名
Journal of Experimental Botany論文タイトル
Improved stomatal opening enhances photosynthetic rate and biomass production in fluctuating light著者
Haruki Kimura, Mimi Hashimoto-Sugimoto, Koh Iba, Ichiro Terashima and Wataru Yamori*DOI番号
https://doi.org/10.1093/jxb/eraa090

用語解説
注1 光合成誘導
光照射後に光合成が始動し、定常状態に達するまでのCO2固定速度の変化過程。この期間は光エネルギーを有効利用することができない。この誘導反応では、暗黒や弱光下で不活性化していた光合成関連酵素が活性化したり、閉じていた気孔が徐々に開くことで光合成の基質であるCO2の供給量が増えたりする。
注2 気孔
葉の表皮に主に存在する開閉式の小さい孔状の構造。孔辺細胞という一対の細胞が膨潤収縮することによって開閉し、植物体と大気間のガス交換(CO2吸収や蒸散)を可能にしている。夜間は閉鎖し不要な蒸散を防ぎ、光照射とともに開口する性質があるほか、湿度やCO2濃度に応じてその開き度合が変化する。
注3 水利用効率
光合成によるCO2同化量を蒸散による水分の損失量で割ることで求められる値。どれだけ水分をうまく利用しつつ光合成できているかの指標の1つである。陸上植物にとって水分の確保は必須であり、この水利用効率の値が高いほど少ない水分で大量のCO2を固定できていることになる。
注4 slac1変異体、ost1変異体
気孔閉鎖経路を欠損したシロイヌナズナ変異体。slac1変異体は陰イオンチャネルを、ost1変異体はプロテインキナーゼを欠損している。結果として、どちらの変異体も気孔を閉鎖すべき環境において経路がうまく働かず、野生型と比較して常に気孔を開いている表現型を示す。
注5 PATROL1過剰発現体
気孔開口に関与するPATROL1タンパク質を過剰発現させたシロイヌナズナ形質転換体。気孔の開口は、孔辺細胞細胞膜上でプロトンATPaseと呼ばれる輸送体が活性化し、プロトン(H+)を放出することでおこる細胞膜電位の過分極(細胞内部が負の膜を隔てた電位差の絶対値が大きくなること)が引き金となって起こる応答である。PATROL1は、気孔を構成する孔辺細胞と隣接する副細胞において、このプロトンATPaseの局在制御に関わるとされる。シロイヌナズナにおいてPATROL1を過剰発現させると、野生型と比較して迅速な気孔開口を示す。
―東京大学大学院理学系研究科・理学部 広報室―

