2022-04-20 理化学研究所
理化学研究所(理研)生命機能科学研究センター比較コネクトミクス研究チームの稲田健吾基礎科学特別研究員、宮道和成チームリーダーらの共同研究グループは、雄マウスが父親となり子育て(養育行動[1])を始める際に鍵となる脳神経回路の変化を明らかにしました。
ライフステージの変化に伴う神経回路の変化を高い解像度で捉えた本研究成果は、父親が示す養育行動の神経基盤を解明する上で重要な知見であり、将来的には人間社会における父親の子育てに関する理解を深めるものと期待できます。
交尾を経験していない雄マウスは仔に対して攻撃的ですが、交尾を経て父親になると、一転して仔を温めたり巣から離れた仔を連れ戻したりするなどの養育行動を示すようになります。
今回、共同研究グループは、視床下部[2]の室傍核[2]と呼ばれる脳領域でオキシトシンを分泌する神経細胞(オキシトシン神経細胞[3])に着目しました。この細胞のオキシトシン合成を阻害すると父親マウスが仔を無視するようになり、逆に神経活動を活性化させると交尾未経験の雄でも養育行動を示したことから、オキシトシン神経細胞が父親マウスの養育行動に必須の役割を持つことが分かりました。さらに、父親マウスの脳内では、オキシトシン神経細胞が活性化しやすくなるような神経入力の変化が起きていることを明らかにしました。
本研究は、科学雑誌『Neuron』(6月15日号)の掲載に先立ち、オンライン版(4月19日付:日本時間4月20日)に掲載されました。

父親となったマウスの脳で起きる神経回路の変化とオキシトシンを介した養育行動の促進
背景
人間社会における父と子の関係は、古来よりさまざまな神話や物語の中でしばしば愛憎劇として描かれてきました。そして近年、父親の育児参加が世界的に重要な社会課題とされており、男性の育児を動物行動学や神経科学など基礎科学の対象として理解する試みが着目されています。しかし、妊娠や出産といった大きなライフイベントを伴う女性と比べ、男性が父親になる過程で、心身にどのような変化が起きるのかについては十分に理解されていません。
行動神経科学の観点から、父親マウスは興味深い研究対象です。交尾未経験の雄マウスは、仔マウスに対して極めて攻撃的です。これは、自分の遺伝子を引き継いでいない他者の仔を排除することで、自分や自分の子孫が生存・繁栄する確率を高めるための行動と考えられています。ところが、交尾や妊娠雌マウスとの同居を経て父親になる時期が近づくと、仔への攻撃性がなくなり、仔を温めたり巣から離れた仔を連れ戻したりするなど、養育行動を示すようになります。このようなライフステージの変化に伴う雄の行動の変化が脳においてどのように制御されているのかは、行動神経科学における未解明の重要な課題です。
そこで、共同研究グループは、脳の神経細胞の接続の仕方そのものが父親マウスにおいて変化しているのではないかと考え、これを検証するための研究を行いました。脳の視床下部と呼ばれる領域には、養育行動に重要であると考えられている神経核[2]が多数存在しています。このうち、室傍核に存在するオキシトシン神経細胞は、マウスを含むげっ歯類の雌において出産や授乳、そして養育行動を促進させる働きを持つ神経ホルモン物質オキシトシンを合成・分泌することが知られています。しかし、雄の養育行動におけるオキシトシンの機能はこれまで良く分かっていませんでした。
研究手法と成果
共同研究グループはオキシトシン神経細胞に着目し、まず、父親の脳内でオキシトシンが分泌されない場合にどうなるのかを調べました。そのために、新しくオキシトシンのコンディショナルノックアウトマウス[4]という特殊なマウス系統を作製しました。このマウス系統では、室傍核のオキシトシン神経細胞に限定して、オキシトシンの合成能を失わせることができます。このコンディショナルノックアウト雄マウスを正常な雌マウスと交尾させ、その後妊娠中から出産5日後までパートナーの雌と同居させることで、「父親マウス」としました。
これまでの研究から、父親マウスは自分の仔でなくても養育行動を示すことが知られており、正常な父親マウスは養育行動を示しました(図1A)。これに対し、オキシトシン合成能を欠損させたコンディショナルノックアウト父親マウスは、養育行動を示す割合が低下し、仔を無視するようになりました(図1A、B)。
交尾未経験の雄マウスは、仔に対して攻撃的です。もしオキシトシンが養育行動を促進するなら、交尾未経験の雄マウスであってもオキシトシン神経細胞を活性化させてオキシトシンを分泌させれば、養育行動を示す可能性があります。そこで次に、薬理遺伝学[5]の手法を使って、交尾未経験の雄マウスのオキシトシン神経細胞を人為的に活性化させました。すると、これらの雄マウスは仔に対して攻撃せず、父親のように養育行動を見せました(図1C、D)。
これらの結果は、オキシトシン神経細胞の活動、そしてオキシトシンの分泌が雄マウスの養育行動を引き起こすために重要であることを示しています。
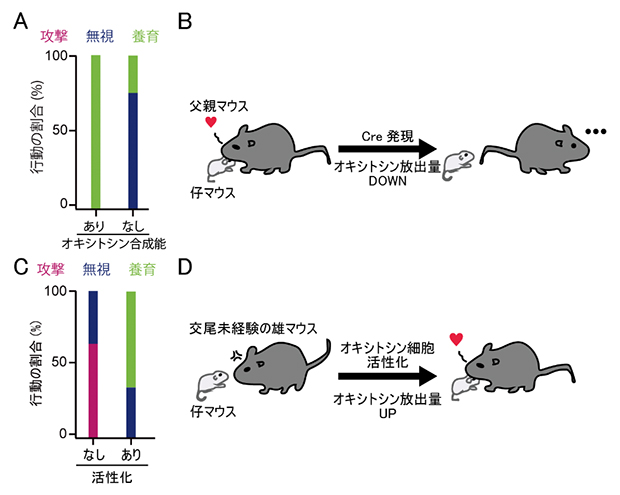
図1 父親マウスと交尾未経験の雄マウスにおけるオキシトシンの効果
A.仔を提示された際の父親マウスの行動の割合。緑は養育行動、青は無視を表す。赤の攻撃行動をとった父親マウスはいなかった。オキシトシン合成能を欠損した父親(右)は、対照群(左)に比べて仔を無視する割合が高い。B.実験結果の模式図。オキシトシンのコンディショナルノックアウト父親マウスは養育行動を示さず、仔を無視するようになる。
C.仔を提示された際に交尾未経験の雄マウスが示した行動の割合。色はAと同じ。
D.実験結果の模式図。交尾未経験の雄マウスのオキシトシン神経細胞を人為的に活性化すると、仔に対する攻撃行動が抑制され、代わりに養育行動が発現する。
これらの実験結果から、「交尾からパートナーの出産に至るまでの過程において、雄マウスの脳内でオキシトシン神経細胞が活性化しやすくなるような神経回路の変化が起きることで、父親マウスの養育行動が支えられているのではないか」との仮説を立てました。そこで、2種類の異なる実験手法を用いて、この仮説を検証しました。
一つ目は、オキシトシン神経細胞に直接入力を送る神経細胞群の特定です。この実験には、狂犬病ウイルスがシナプス後細胞からシナプス前細胞へと広がる性質を持つことを利用したトランスシナプス標識法[6]を用いました(図2A)。交尾未経験雄マウスと父親マウスのオキシトシン神経細胞のシナプス前細胞群を網羅的に比較解析した(図2B)結果、外側視床下部と呼ばれる脳領域からの入力が父親では著しく増えていることが分かりました。さらに、このシナプス前細胞が興奮性か抑制性か調べたところ、興奮性の入力が増えていることが分かりました(図2C)。
二つ目は、オキシトシン神経細胞に入力される神経伝達強度の解析です(図2D)。この実験では、光遺伝学[7]と電気生理学的手法[8]を用いました。光遺伝学は、光に反応するタンパク質を標的となる神経細胞に発現させ、光を当てている間だけその細胞からの神経伝達物質の放出を誘起させる手法です。本実験では、オキシトシン神経細胞に入力する外側視床下部の興奮性神経細胞を標的としました。一方、オキシトシン神経細胞からは、全細胞記録法という手法を用いて、神経細胞の電気活動を高精度で記録しました。その結果、外側視床下部からの興奮性入力が、父親マウスの方が交尾未経験の雄マウスよりも強いことが分かりました(図2E、F)。
これら二つの独立した実験により、外側視床下部興奮性神経細胞からオキシトシン神経細胞への入力強度が、父親において増強されていることが分かりました。
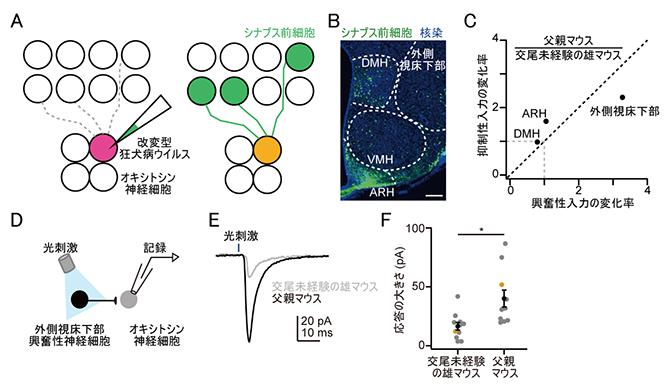
図2 父親マウスと交尾未経験の雄マウスにおけるオキシトシン神経細胞への入力強度の比較
A.トランスシナプス標識法の模式図。(左)オキシトシン神経細胞の一部に遺伝学的な操作(赤色蛍光タンパク質による細胞標識)を施した後、緑色蛍光タンパク質遺伝子を組み込んだ改変型狂犬病ウイルス(感染能力を限定し無毒化したウイルス)を感染させる。(右)改変型狂犬病ウイルスは緑色蛍光タンパク質を発現するため、一次感染したオキシトシン神経細胞は黄色(赤色と緑色の加色)に標識される。一方、黄色の神経細胞に対してシナプス入力を形成しているシナプス前細胞は、緑色で網羅的に可視化される。
B.トランスシナプス標識法を施した脳切片の染色図。オキシトシン神経細胞に対してシナプス入力を形成しているシナプス前細胞(緑色)の神経核の名前を白色の文字で示す。DMH;背内側核、VMH;腹内側核、ARH;弓状核。スケールバーは100マイクロメートル(μm、1μmは1,000分の1mm)。
C.一つのオキシトシン神経細胞への興奮性および抑制性の入力細胞数の変化率。父親マウスのオキシトシン神経細胞への入力細胞数が交尾未経験の雄マウスと同じ場合は1、父親の方が多い場合1より大きくなる。父親マウスにおいて、背内側核(DMH)は変化が見られず、弓状核(ARH)では抑制入力がやや増えた一方、外側視床下部の興奮性神経細胞からオキシトシン神経細胞への入力が著しく増えている。
D.光遺伝学と電気生理学的手法の模式図。外側視床下部の興奮性の神経細胞に光感受性タンパク質を発現させ、青色光で刺激すると、シナプスから神経伝達物質が放出される。この入力を受けたオキシトシン神経細胞からは、電気生理学的手法で電気活動を記録する。
E.父親マウスと交尾未経験の雄マウスの細胞応答。父親の応答の方が大きい。L字型のスケールは、細胞応答の大きさを示す縦軸の電流変化(pA;ピコアンペア)と時間の幅(ms;ミリ秒)を示す。
F.父親マウスと交尾未経験の雄マウスの細胞応答の大きさの定量解析。点はそれぞれのマウスで電気活動を記録した細胞のデータ点を示し、黄色点はEで示したデータ点。黒点はそれぞれの平均値。*比較した2群の有意水準;p < 0.05。
では、外側視床下部の興奮性神経細胞からオキシトシン神経細胞への入力強度が、父親で増えていることには、どのような意味があるのでしょうか。外側視床下部は、これまで養育行動の研究ではあまり注目されてきませんでした。そこで、まず、父親マウスの外側視床下部が仔との触れ合いで活性化されるかを、神経活動の指標となる遺伝子c-fosの発現により調べました。解析の結果、父親マウスの外側視床下部の神経細胞は仔との触れ合いで活性化されており(図3A)、またその多くが興奮性神経細胞であることも分かりました(図3B)。
最後に、交尾未経験の雄マウスにおいて、外側視床下部の興奮性神経細胞を人為的に活性化した際の、仔への行動を調べました。すると、養育行動こそ出ないものの、仔への攻撃行動が抑制されました(図3C)。この攻撃行動の抑制はオキシトシンのノックアウトマウスでは起こらなかったことから、外側視床下部は下流のオキシトシン神経細胞の活性化を介して仔への攻撃性を下げる役割を果たすことを示しています。一方で、外側視床下部の活性化だけでは養育行動を生み出せないことから、他の神経核からオキシトシン神経細胞へのさまざまな入力が協調的に働くことが、父親の養育行動に必要である可能性が示されました(図3D)。
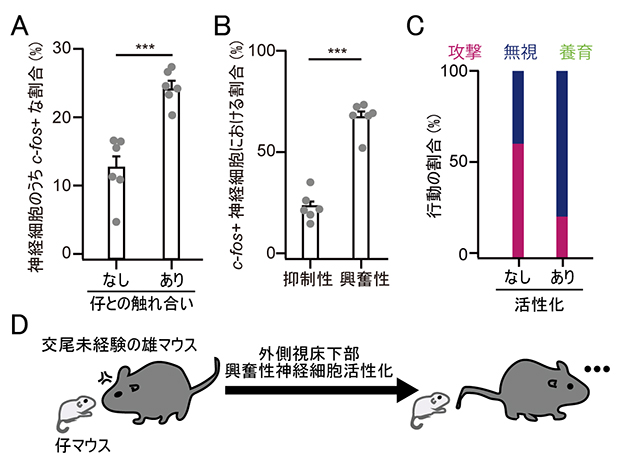
図3 仔との触れ合いで活性化される外側視床下部の興奮性神経細胞
A.父親マウスの外側視床下部において、神経細胞が活性化されたかどうかのマーカーとしてc-fosを染色し、c-fosを発現する細胞(c-fos+)の割合を計測した。外側視床下部の神経細胞は、仔と触れ合った父親マウスの方がより活性化される(***比較した2群の有意水準;p < 0.001)。
B.Aで調べた、活性化された細胞の多くは興奮性の細胞である(***比較した2群の有意水準;p < 0.001)。
C.交尾未経験の雄マウスにおいて外側視床下部の興奮性神経細胞を人為的に活性化させると、養育行動は誘導されなかったが、攻撃行動が抑制された。
D.実験結果の模式図。外側視床下部の興奮性神経細胞は、雄マウスにおいて仔への攻撃性を抑える役割がある。
今後の期待
今回共同研究グループは、交尾から出産までパートナーの雌マウスと連れ添うことで、父親マウスの脳内、特にオキシトシン神経細胞への神経入力が変化することを明らかにしました。しかし、交尾や雌マウスとの同居において、具体的にどのような刺激がこの変化の引き金になっているのかは不明であり、また神経回路の変化の背景にある分子・神経メカニズムも分かっていません。今回、ライフステージの変化に伴う神経回路レベルの変化が実際に見つかったことで、オキシトシン神経細胞や他の細胞群への神経配線も含めたより包括的な研究が必要になると考えられます。
今後さらに研究を進め、神経回路の変化を引き起こす要因の解明やその進化的保存性の検討などを通して、ヒトを含む哺乳類の養育行動を支える神経基盤が明らかになるものと期待できます。
補足説明
1.養育行動
仔を保護し養育する行動の総称。本研究では、特に仔をなめて清潔に保つグルーミング行動や、巣の外にいる仔を安全な巣の中に連れ帰る行動、そして巣の中で仔を保温する行動を主な解析対象とした。なお、父親になる時期が近づくと何らかの養育行動が雄に発現する現象は、マウス以外にも多くの動物で観察されている。
2.視床下部、室傍核、神経核
視床下部は、内分泌や自律機能の調節を担い生理機能をつかさどる中枢領域。神経核は、中枢神経において神経細胞の細胞体が塊状に存在している領域を指す用語。視床下部は、特定の機能をつかさどる数多くの神経核から構成されており、本研究で扱った室傍核や外側視床下部も視床下部に含まれる神経核である。室傍核は神経内分泌の中枢の一つとして知られ、ストレス応答や代謝制御に関与するホルモンの分泌を促進する機能の他、オキシトシンや抗利尿ホルモン(バソプレシン)を分泌する神経細胞がこの神経核に含まれる。外側核は、睡眠や代謝、記憶の制御、摂食行動との関連が指摘されているが、養育行動への関与についてはこれまでほとんど知見が無かった。
3.オキシトシン神経細胞
細胞内で合成した神経ホルモン物質オキシトシンを分泌する神経細胞。視床下部では主に室傍核と視索上核という二つの神経核に局在している。オキシトシン神経細胞が、脳下垂体後葉を経由して血中に分泌するオキシトシンは、雌の出産や射乳に重要であることが知られており、ヒトではオキシトシン製剤が陣痛促進剤として使用されている。オキシトシン神経細胞は血中だけでなく、脳内にもオキシトシンを分泌する。本研究では、この脳内に分泌されるオキシトシンに注目している。
4.オキシトシンのコンディショナルノックアウトマウス
細胞内におけるオキシトシンの産生に必要なDNA配列を、DNA組換え酵素Cre依存的に欠損させることのできるマウス系統。「Creを発現している細胞だけにおいて」という条件付き(コンディショナル)で作動するので、従来のオキシトシンノックアウトマウスではできなかった時間的・空間的な制御が可能。本研究ではCreを発現させるアデノ随伴ウイルス(AAV)を左右の室傍核に局所注入することで、室傍核のオキシトシン神経細胞からオキシトシンの合成能を欠損させた。
5.薬理遺伝学
薬理遺伝学では、内在性のリガンドに応答せず特定の薬剤のみに応答するように改変した人工的な受容体を、分子遺伝学を用いて標的の神経細胞群に発現させた後、その薬剤を投与することで、神経細胞を人為的に興奮させたり、抑制させたりする。本研究では、DREADDと呼ばれる人工的な受容体を使用して、目的の細胞の神経活動を操作するのに用いた。
6.トランスシナプス標識法
ある神経細胞群へシナプス入力を送るシナプス前細胞を網羅的に可視化する手法。本研究では、室傍核のオキシトシン神経細胞のシナプス前細胞を可視化した。本手法で用いた改変型狂犬病ウイルスは、(i)TVA受容体という哺乳類が本来持っていない受容体を発現する細胞にだけ感染する、(ii)感染後シナプス前細胞に感染を広げるために必須なタンパク質RG(rabies glycoprotein)を作れない、そして(iii)感染した後に緑色蛍光タンパク質を発現する、という改変を施されている。本研究では、オキシトシン神経細胞にTVA受容体とRGを発現させた上で、この改変型狂犬病ウイルスを感染させることで、オキシトシン神経細胞のシナプス前細胞を緑色蛍光タンパク質で標識・解析した。
7.光遺伝学
光遺伝学では、特定の波長の光に反応してイオンを輸送する光感受性タンパク質を、分子遺伝学を用いて特定の神経細胞群に発現させた後、その神経細胞群に光を当てることで、神経細胞を興奮させたり、抑制させたりする。本研究では、青色光に反応して神経細胞を興奮させるチャネルロドプシン2を使用した。
8.電気生理学的手法
神経細胞の電気活動を記録する実験手法の総称。本研究では、全細胞記録(ホールセル)法という手法を用いた。この手法では、ガラスピペットの中に電極を入れ、そのガラスピペットをギガ(10億)Ω以上の高抵抗で細胞体表面に密着させた後、わずかに穴を開ける。これによりノイズを減らすことができ、神経細胞内の微弱な電気活動も精密に記録できる。
共同研究グループ
理化学研究所 生命機能科学研究センター
比較コネクトミクス研究チーム
チームリーダー 宮道 和成(みやみち かずなり)
基礎科学特別研究員 稲田 健吾(いなだ けんご)
テクニカルスタッフ 萩原 光恵(はぎはら みつえ)
テクニカルスタッフ(研究当時) 辻本 和子(つじもと かずこ)
生体モデル開発チーム
チームリーダー 清成 寛(きよなり ひろし)
技師 阿部 高也(あべ たかや)
群馬大学 医学部
教授 平井 宏和(ひらい ひろかず)
講師 今野 歩(こんの あゆむ)
研究支援
本研究は、理化学研究所運営費交付金(生命機能科学研究)で実施し、日本学術振興会(JSPS)若手研究「オキシトシンによる攻撃性緩和の神経回路機構(研究代表者:稲田健吾)」、同特別研究員奨励費「神経配線のレベルにおける雌雄差の形成メカニズム(特別研究員:稲田健吾)、同基盤研究(B)「比較コネクトミクスの創出(研究代表者:宮道和成)」、花王芸術・科学財団花王科学奨励賞(稲田健吾)、科学技術振興機構(JST)さきがけ(宮道和成)などによる支援を受けて行われました。
原論文情報
Kengo Inada, Mitsue Hagihara, Kazuko Tsujimoto, Takaya Abe, Ayumu Konno, Hirokazu Hirai, Hiroshi Kiyonari & Kazunari Miyamichi, “Plasticity of neural connections underlying oxytocin-mediated parental behaviors of male mice”, Neuron, 10.1016/j.neuron.2022.03.033
発表者
理化学研究所
生命機能科学研究センター 比較コネクトミクス研究チーム
チームリーダー 宮道 和成(みやみち かずなり)
基礎科学特別研究員 稲田 健吾(いなだ けんご)
報道担当
理化学研究所 広報室 報道担当
の構造を世界で初めて決定~サポウイルスの治療薬やワクチンの開発を加速~")
では赤ちゃんが小さく生まれる」の謎を解明")